![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 39 (Nº 37) Ano 2018 • Página 28
Denise de Andrade CUNHA 1; Leandro Valle FERREIRA 2;
Recebido: 01/04/2018 • Aprovado: 15/05/2018
RESUMO: O objetivo desse trabalho foi analisar a dinâmica do incremento e perda em biomassa de populações de Virola surinamensis (Rol.) Warb. em função da razão sexual, nas florestas de várzea da Estação Científica Ferreira Penna, Brasil. Os resultados sugerem que os individuos femininos contribuem com maior tempo de residência de carbono fixado na biomassa viva. Destacamos assim, a importância dos indivíduos femininos de V.surinamensis não apenas para conservação da espécie como importantes para o estoque de carbono nessas florestas. |
ABSTRACT: The objective of this work was to analyze the dynamics of biomass increase and loss of populations of Virola surinamensis (Ro.) Warb. depending on the sex ratio in white-water forests of the Ferreira Penna Research Station, Brazil. The results suggest that female individuals contribute with a longer residence time of carbon fixed in living biomass. We emphasize the importance of V.surinamensis female individuals not only for the conservation of the species as important for the carbon stock in these forests. |

Estudos indicam que a grande capacidade de acumular carbono que as florestas tropicais apresentam é resultado da sua grande produtividade primária líquida, a qual pode ser definida como a quantidade líquida de carbono que é assimilado da atmosfera e fixado em nova matéria orgânica por unidade de tempo (Malhi et al., 2002; Philips et al., 2009). Desse modo, a produtividade pode ser estimada através de modelos que utilizam parâmetros fisiológicos e/ou estimando o acúmulo de biomassa ao longo do tempo, o qual pode ser efetuado através de equações alométricas (Higuchi et al., 2006).
O acúmulo em biomassa está diretamente relacionado com a eficiência no sequestro de carbono pelos vegetais (Malhi et al., 2004; Malhi, et al., 2006). De modo complementar, o tempo de residência de carbono, que corresponde ao tempo em que o carbono permanece fixado na biomassa lenhosa viva acima do solo (Malhi et al., 2004), é reflexo da dinâmica florestal e da sua eficácia como sumidouro de carbono.
Na Amazônia, a maioria absoluta de estudos que relacionam produtividade e dinâmica florestal foram desenvolvidos em florestas de terra firme. Pouco se conhece a respeito da capacidade das florestas inundáveis da Amazônia em acumular biomassa ao longo do tempo e sua, consequente, eficiência em estocar carbono (e.g. Xavier, 2009)
As florestas inundáveis possuem grande representatividade ocupando, aproximadamente, 14% da bacia amazônica (Melack & Hess, 2010). Grande parte dessas florestas inundáveis são caracterizadas como ‘várzeas’ que, ocupando aproximadamente 400.000 km2 da bacia Amazônica (Melack & Hess, 2010), apresentam-se associadas a rios que possuem pH neutro e altas concentrações de nutrientes dissolvidos, originados dos Andes e de outras formações recentes, o que resulta em áreas de elevada fertilidade (Furch & Junk, 1997; Furch, 1997; Worbes, 1997; Junk et al., 2012).
Em função desse aporte de nutrientes, estudos apontam alta produtividade em florestas de várzea (Malhi et al., 2004) já que os indivíduos arbóreos apresentam elevada biomassa devido seu rápido crescimento (Worbes, 1997; Schöngart et al., 2005).
O funcionamento do ecossistema, no que diz respeito, por exemplo, à produtividade, é regulado, principalmente, por características das espécies mais abundantes (Grime, 1998; Garnier et al., 2004; Diaz et al., 2007). Nesse sentindo, as espécies mais abundantes podem controlar, em grande parte, o ganho e a perda de biomassa em florestas.
Entretanto, as florestas de várzea, têm sido historicamente, muito utilizadas para atividades econômicas de extrativismo vegetal onde o padrão da exploração madeireira está associado a uma alta concentração de exploração de poucas espécies de alto valor comercial o que tem diminuindo os estoques naturais destas espécies (Junk, 2000) que outrora apresentavam grande abundância.
Nesse contexto, Virola surinamensis (Rol.) Warb., uma espécie dioica típica de florestas inundáveis e de ampla distribuição na Amazônia, apesar de apresentar alta abundância em florestas de várzea de áreas protegidas (Ferreira et al., 2010; Ferreira et al., 2014), encontra-se na lista vermelha de espécies ameaçadas de extinção (Americas Regional Workshop, 1998).
Atualmente, a maioria das florestas de várzeas do estuário amazônico tem escassos estoques de V. surinamensis (Salomao et al., 2007; Ferreira et al., 2014) devido a intensa exploração madeireira concentrada, principalmente, na região do estuário amazônico, onde se encontram grandes pólos da atividade madeireira (Piña-Rodrigues & Mota, 2000, Salomão et al., 2007).
Faz-se necessário, portanto, o estudo da capacidade em armazenar carbono de populações em florestas de várzea a partir do monitoramento dos padrões de produtividade e dinâmica desses indivíduos cujo estudo baseia-se na quantificação do ganho, através do incremento, e perda, por mortalidade, em biomassa lenhosa acima do solo.
O objetivo desse trabalho foi analisar a dinâmica da biomassa lenhosa acima do solo a partir de dados de incremento e mortalidade de populações de V. surinamensis em função da razão sexual em florestas de várzea da Estação Científica Ferreira Penna-Caxiuanã, Pará.
O estudo foi realizado em florestas de várzea na área da Estação Científica Ferreira Penna (ECFPn) situada no território da Floresta Nacional (FLONA) de Caxiuanã no município de Melgaço no Estado do Pará, a cerca de 400 km de Belém, nas proximidades da Baía de Caxiuanã, entres os rios Xingu e Tapajós (Figura 1).
Figura 1
Localização da Floresta Nacional de Caxiuanã no estado do Pará (A),
posição da Estação Científica Ferreira Penna em relação à Floresta Nacional
de Caxiuanã (B) e os tipos de vegetações da Estação Científica Ferreira Penna (C).
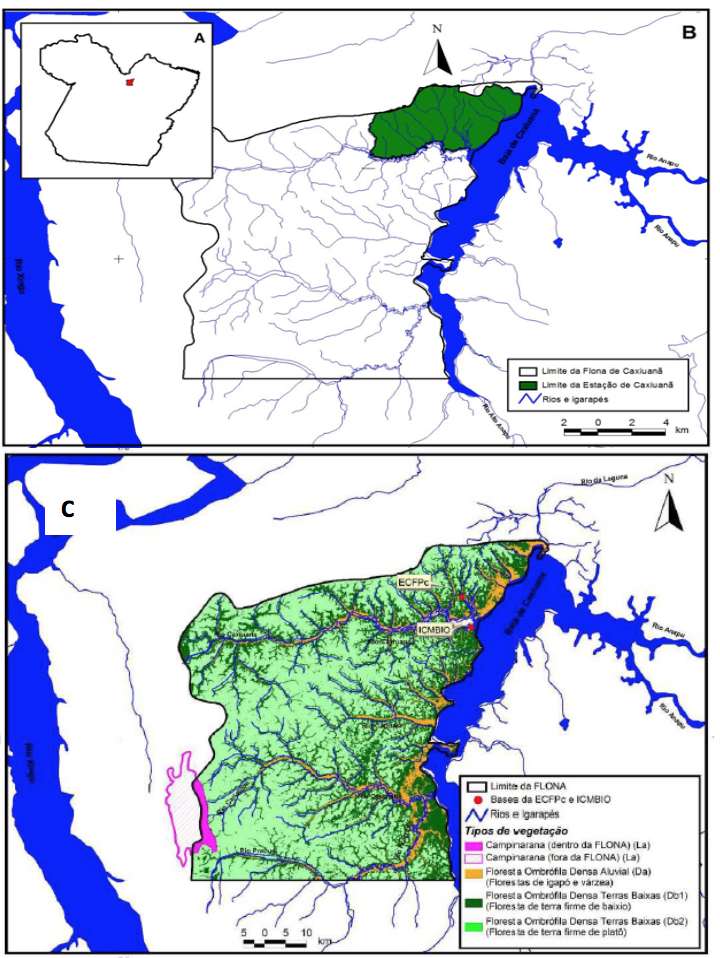
As florestas de várzea inventariadas neste estudo estão localizadas às margens da Baía de Caxiuanã a qual apresenta cerca de 40 km de comprimento e 8-15 km de largura (Behling, 2011), pH neutro e carga de nutrientes elevada (Icmbio, 2012). Essa grande quantidade de nutrientes é resultado da alta carga de sedimentos dissolvidos oriundos dos Andes que chegaram à Baía de Caxiuanã devido uma ligação com o Rio Amazonas que foi perdida em decorrência da transformação do rio Anapu ocorridas durante as últimas transgressões marinhas (Berredo et al., 2012).
A vegetação é caracterizada pela elevada área basal (Ferreira et al., 2005) e grande domínio de indivíduos de Euterpe oleracea Mart., Pterocarpus santalinoides L'Hér. ex DC., Virola surinamensis e Pentaclethra macroloba (Willd.) Kuntze (Ferreira et al., 2013).
Em 2011, foram instaladas seis parcelas permanentes de 20 x 100 m (0,2 ha) dispostas paralelamente às margens da Baía de Caxiuanã para o monitoramento da vegetação.
Em cada parcela, todas as V. surinamensis e demais indivíduos com diâmetro a altura do peito (DAP) ≥ 10 cm foram identificados, medidos os DAP’s e marcados com placas de alumínio.
O acompanhamento fenológico dos indivíduos de V. surinamensis marcados no censo inicial de 2011 foi realizado mensalmente durante os anos de 2012 e 2013.
A remedição do DAP dos indivíduos foi feita anualmente no mês de dezembro em todas as parcelas permanentes até o ano de 2017. Durante as remedições anuais, as medidas foram supervisionadas para que o diâmetro fosse medido exatamente no mesmo local da medida inicial.
A mortalidade foi considerada como o número de árvores mortas em cada censo, incluindo aquelas encontradas em pé e aquelas caídas no chão.

A pressuposição de normalidade dos dados foi testada usando o teste de Shapiro-Wilk e a homogeneidade das variâncias foi avaliada pelo teste de Levene (Shapiro & Wilk, 1965; Zar, 2010).
O Test t de Student com nível de significância de 5% (Zar, 2010)para testar as diferenças no número de individuos por sexo entre as parcelas.
As diferenças das médias anuais de biomassa dos individuos, incremento em biomassa e biomassa perdida por mortalidade (variáveis dependentes) em função da razão sexual (fatores) foram testadas utilizando o Test t de Student com nível de significância de 5% (Zar, 2010).
Foi utilizada a análise de variância simples (ANOVA) para medidas repetidas para analisar as diferenças da biomassa por parcela (variávies dependentes) entre os anos inventariados (fatores) e também para verificar o número de indivíduos e biomassa (variável dependente) entre as classes de diâmetro (10–19.9, 20–29.9, 30–49.99, ≥ 50cm) (fatores) em função do sexo de V.surinamensis (Zar, 2010).
Nas seis parcelas analisadas foram registrados 1.116 indivíduos distribuídos em 45 espécies, entre as quais V. surinamensis foi a mais abundante com média anual de 23,3% dos indivíduos amostrados.
Foram registrados 120 indivíduos femininos, 173 indivíduos masculinos e 23 indivíduos que não apresentaram período reprodutivo de V.surinamensis. A biomassa total média desses indivíduos foi de 187,1 Mg.ano-1 (Tabela 1).
Tabela 1
Média da biomassa total acumulada, o incremento em biomassa, número de
mortos e biomassa perdida por mortalidade entre os indivíduos do sexo feminino e
masculinos de Virola surinamensis (Rol.) Warb..
Sexo |
Média da biomassa acumulada (Mg.ano-1) |
Média do incremento em biomassa (Mg.ano-1) |
Nº de mortos |
Biomassa perdida por mortalidade (Mg.ano-1) |
Feminino |
95,7 |
1,2 |
11 |
37,1 |
Masculino |
88,4 |
1,9 |
19 |
16,4 |
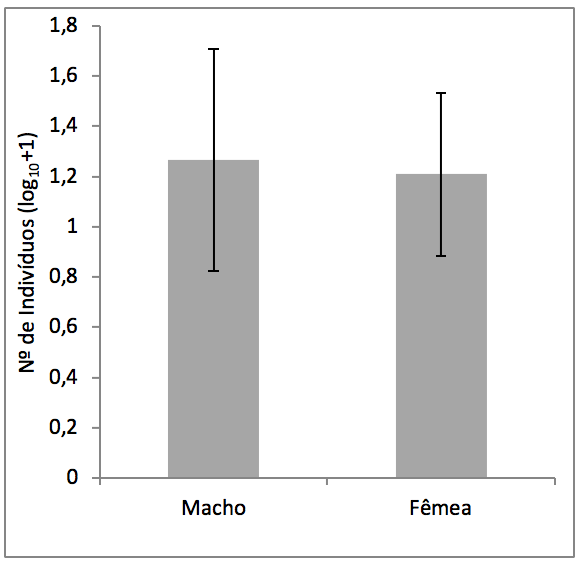
Não houve diferença significativa no número de indivíduos entre os sexos (t = 0,32; p = 0,37) (Figura 2). Contudo, os indivíduos femininos apresentaram biomassa significativamente maior que os indivíduos masculinos (16,02 ± 8 vs. 12,6 ± 9 Mg/0,2ha/ano; F = 4,34; p= 0,03). Essas diferenças porém não foram significativas ao longo dos anos (F = 0,0002; p = 1).
Figura 2
Média e desvio padrão da número de indivíduos (log10+1)
de fêmeas e machos de Virola surinamensis (Rol.) Warb.
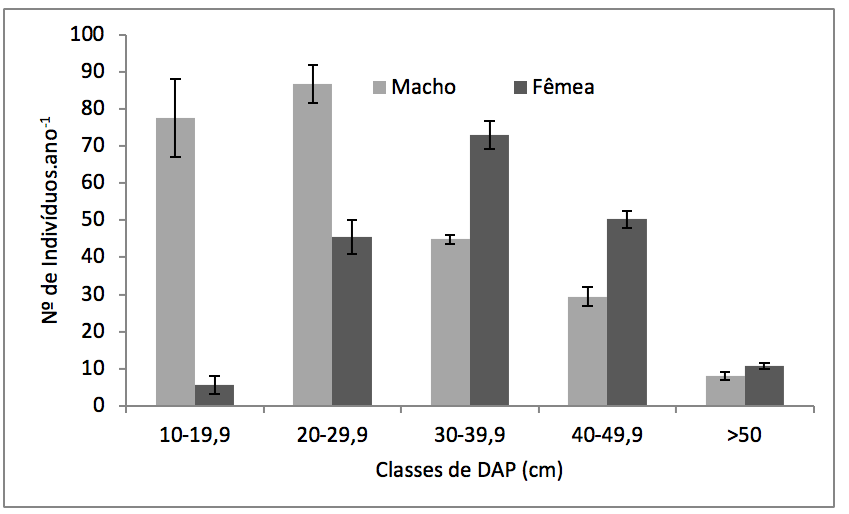
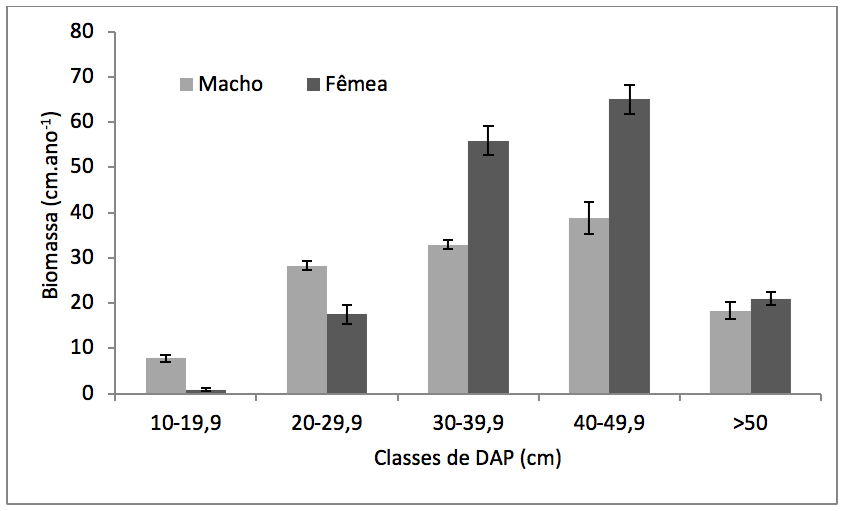
Houve diferença significativa no número de machos e fêmeas por classe de DAP (F= 348,77; p< 0,0001). Os machos apresentaram maior número de indivíduos nas primeiras classes de DAP (10cm à 29,9cm), enquanto que as fêmeas apresentaram maior número de indivíduos nas classes de DAP de 30cm à 49,9cm (Figura 3). Com relação à biomassa, houve maior acúmulo de biomassa nas clases superiores à 30 cm de DAP pelos individuos femininos (F = 220,53; p < 0,0001) (Figura 4).
Figura 3
Média e desvio padrão anual do número de indivíduos de Virola surinamensis (Rol.)
Warb. masculinos e femininos por classe de diâmetro à altura do peito (DAP).
------
Figura 4
Média e desvio padrão anual da biomassa de Virola surinamensis (Rol.) Warb.
masculinos e femininos por classe de diâmetro à altura do peito (DAP).
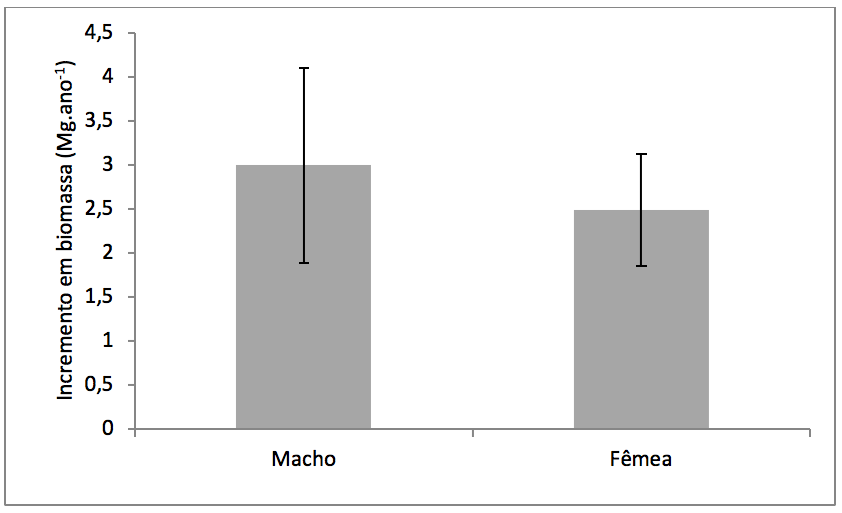
Não houve diferença no incremento em biomassa entre os sexos de V. surinamensis (Macho: 2,9 ± 1,1; Fêmea: 2,4 ± 0,6 Mg.ano-1; t = 0,9; p= 0,1) (Figura 5).
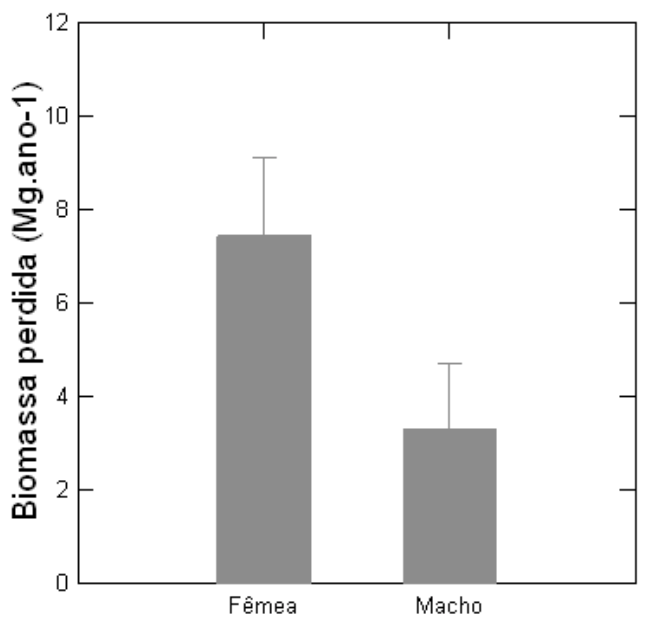
Foram registrados 34 indivíduos mortos, dos quais 11 eram fêmeas e 19 eram machos. Contudo, a perda de biomassa por mortalidade foi significativamente maior entre os indivíduos femininos de V. surinamensis quando comparadas com os indivíduos masculinos (7,4 ± 3,3 vs. 3,2 ± 2 Mg.ano-1; t = 2,1; p = 0,05) (Figura 6).
Figura 5
Média e desvio padrão do incremento em biomassa anual de indivíduos
femininos e masculinos de Virola surinamensis (Rol.) Warb.
-----
Figura 6
Média e desvio padrão da biomassa perdida por mortalidade entre indivíduos
masculinos e femininos de Virola surinamensis (Rol.) Warb.
As médias de abundâncias estatisticamente semelhantes entre indivíduos masculinos e femininos estão em concordância quanto à razão sexual de 1:1 esperada para espécies dioicas de florestas tropicais (Opler & Bawa, 1978).
Contudo, houve menor número de indivíduos femininos nas primeiras classes de DAP (10cm-19,9cm; 20cm-29,9cm) o que também foi registrado por Ferreira et al. (2014) nas florestas de várzea de Caxiuanã. Este resultado pode sugerir, aparentemente, que a população feminina não está em equilíbrio já que para Kurtz & Araujo (2000), quando a série de classes de diâmetros é interrompida ou truncada em qualquer um dos extremos, o ciclo de vida das plantas não está se completando e a população geralmente não pode ser considerada em equilíbrio.
Entretanto, Opler & Bawa (1978), em estudo realizado em espécies dioicas de florestas tropicais, sugerem que indivíduos masculinos possuem maturação sexual significativamente diferente dos femininos.
Piña-Rodrigues (1999) afirma que V.surinamensis apresenta comportamento diferencial em relação à idade inicial de florescimento, com alguns indivíduos se destacando por sua precocidade.
Isso pode ocorrer porque a alocação diferencial de recursos e os diferentes gastos energéticos interferem na idade reprodutiva entre indivíduos de espécies dioicas: as plantas femininas, devido sua maior necessidade de alocar reservas para suprir a grande demanda energética durante o período reprodutivo, alcançam tardiamente a idade reprodutiva quando comparadas com os indivíduos masculinos (Armstrong & Irvine, 1989).
No caso de V. surinamensis, a maturação sexual de indivíduos femininos pode estar fortemente relacionada com indivíduos acima de 30 cm de DAP. Esse resultado tem importantes implicações para a conservação, já que o equilíbrio populacional dessa espécie pode estar relacionado com manutenção desses indivíduos nas florestas de várzea. Nesse sentido, a Instrução Normativa 40/2010 da SEMA/PA demonstra ser adequada para V. surinamensis já que estabelece o limite mínimo de corte seletivo de 50 cm de diâmetro para indivíduos de espécies madeireiras nas florestas de várzea (Ferreira et al., 2014). Para isso, seria necessário um tempo superior à 40 anos já que Piña-Rodrigues (1996) relatou que, em plantios no estuário, V.surinamensis só atingiu DAP > 40 cm entre 30 – 40 anos.
O incremento periódico anual em biomassa diz respeito ao ganho em biomassa a partir do crescimento em DAP dos indivíduos de V. surinamensis o qual não apresentou diferença significativa entre indivíduos femininos e masculinos sugerindo que, apesar das diferenças intrasexuais em estratégias reprodutivas (Piña-Rodrigues, 1999; Lenza & Oliveira, 2006), esses indivíduos apresentem semelhantes estratégias de alocação de recurso para crescimento em diâmetro.
Essa característica pode ser resultado das pressões seletivas impostas, principalmente, pelo constante estresse hídrico por inundação às populações de florestas de várzea resultando em baixa diversidade intraespecífica (Ferreira et al., 2013).
A maior mortalidade apresentada por indivíduos masculinos pode estar relacionada com características como a altura e herbivoria, as quais não foram analisadas nesse estudo. A altura da planta, por exemplo, está relacionada com sua capacidade competitiva de adquirir luz para fotossíntese (Sumida et al., 1997). Contudo, grandes taxas de crescimento em altura podem ser obtidas às custas de crescimento em diâmetro necessário para suporte mecânico e fisiológico da planta (King, 1991). Assim, árvores que investem menos em apoio estrutural podem crescer mais rapidamente, porém, podem apresentar menor resistência mecânica e sobrevivência (King et al., 2006).
A herbivoria também pode influenciar na mortalidade de indivíduos masculinos já que o custo energético para produção de galhas e os danos causados às flores exercem elevado gasto para as plantas masculinas, resultando, com o passar do tempo, na morte desses indivíduos (Wolf, 1997).
A não diferença nas médias das abundâncias associada a não diferença nas médias de incremento anual em biomassa entre indivíduos femininos e masculinos ao longo dos anos demonstram que ambos os sexos contribuem de forma similar na produtividade primária líquida das florestas de várzea. A produtividade primária líquida é definida como a quantidade líquida de biomassa fixada em matéria orgânica por unidade de tempo (MALHI et al., 2002).
Contudo, mesmo com alta biomassa perdida por mortalidade, os indivíduos femininos, por um lado, apresentaram menor mortalidade e, por outro, possuíram maior biomassa total acumulada ao longo dos anos o que pode ser explicado pelo maior número desses indivíduos nas maiores classes de DAP em detrimento dos indivíduos masculinos que se apresentaram em menor número. Esses resultados demonstram maior eficácia dos indivíduos femininos no estoque de carbono já que parecem contribuir com maior tempo de residência de carbono fixado na biomassa viva.
Essa característica é muito importante para florestas de várzea as quais, devido sua expressiva produtividade e alta dinâmica, podem não apresentar função significativa como sumidouro de carbono já que as altas taxas de crescimento de indivíduos podem não compensar o menor tempo de permanência do carbono na biomassa devido a altas taxa de mortalidade nesses ambientes. Malhi et al. (2004) sugere que nos trópicos o tempo de residência seja de 20 em floretas com alta produtividade. Adicionalmente, florestas maduras de várzea com mais de 200 anos podem apresentar um equilíbrio no balanço entre ganhos e perdas de biomassa indicando que, nessas florestas, a biomassa não tem função significativa como sumidouro de carbono (Xavier, 2009).
Assim, levando em consideração a alta dinâmica atribuída às florestas de várzea e grande abundância de V. surinamensis, ressaltamos a importância dos indivíduos femininos no prolongamento do tempo de residência de carbono na biomassa lenhosa acima do solo nessas florestas.
Os indivíduos femininos e masculinos de V. surinamensis não apresentaram diferenças significativas na abundância e incremento em biomassa. Contudo, o maior número de indivíduos femininos concentrados nas maiores classes de DAP, menor mortalidade e consequente maior acúmulo de biomassa total, sugerem que estes indivíduos contribuem com maior tempo de residência de carbono fixado na biomassa viva. Destacamos aqui, portanto, a importância dos indivíduos femininos de V. surinamensis não apenas para conservação da espécie como eficaz no estoque de carbono.
Americas Regional Workshop (Conservation & Sustainable Management of Trees, Costa Rica, November 1996). (1998). Virola surinamensis. The IUCN Red List of Threatened Species 1998: e.T33959A9816820. Recuperado de http://dx.doi.org/10.2305/IUCN.UK.1998.RLTS.T33959A9816820.en.
Armstrong, J.E., y Irvine, A.K. (1989). Flowering, sex ratio, pollen-ovule ratios, fruit set and reproductive effort of a dioecious tree, Myristica insipida (Myristicaceae), in two different rain forest communities. American Journal of Botany, 76, 74-85.
Baker, P.J., Phillips, O.L., Malhi, Y., Almeida, S., Arroyo, L., Di Fiori, A., Erwin, T., Killeen, T.J., Laurance, S.G., Laurance, W.F., Lewis, S.l., Lloyd, J., Monteagudo, A., Neill, D.A., Patiño, S., Pitman, N.C.A., Silva, J.N.M. y Martínez, R.V.( 2004). Variation in wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology, 10, 545-562.
Behling, H. (2011). Holocene environmental dynamics in coastal, eastern and central Amazonia and the role of the Atlantic sea-level change. Geographica Helvetica, 66(3), 208-216.
Berredo, J.F., Melo, D.M.B., Ribeiro, M.M., Migiyama, A.C., y Sales, M.E. (2012). Hidrografia da Floresta Nacional de Caxiuanã. En: Instituto Chico Mendes de Conservação da Biodiversidade (eds.), Plano de Manejo da Floresta Nacional de Caxiuanã (pp. 128-132). ICMbio, Brasília.
Diaz, S., Lavorel, S., De Bello, F., Quétier, F., Grigulis, K., y Robson, T. M. (2007).
Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Science, 104, 20684-20689.
Ferreira, F.V., Almeida, S.S., Amaral, D.D., y Parolin, P. (2005). Riqueza e composição de espécies da floresta de igapó e várzea da Estação Científica Ferreira Penna: subsídios para o plano de manejo da Floresta Nacional de Caxiuanã. Pesquisas Botânica, 56, 103-116.
Ferreira, L.V., Almeida, S.S., y Parolin, P. (2010). Amazonian white- and blackwater floodplain forests in Brazil: large differences on a small scale. Ecotropica, 16, 31–41.
Ferreira, L.V., Chaves, P.P., Cunha, D.A., Matos, D.C.L., y Parolin, P. (2013). Variação da riqueza e composição de espécies da comunidade de plantas entre as florestas de igapós e várzeas na Estação Científica Ferreira Penna-Caxiuanã na Amazônia Oriental. Pesquisas Botânica, 64, 175-195.
Ferreira, L.V., Cunha, D.A., y Parolin, P. (2014). Effects of logging on Virola surinamensis in an Amazonian floodplain forest. Environment Conservation Journal, 15(3), 1-8.
FINGER, C.A.G., 1992. Fundamentos de biometria florestal. Santa Maria. 269p.
Furch, K. (1997). Chemistry of várzea and igapó soils and nutrient inventory of their floodplain forests. En, W.J. Junk (eds.), The Central Amazon Floodplains: Ecology of a Pulsing System (pp. 47-67). Berlin: Springer Verlag.
Furch, K., y Junk, W.J. (1997). Physicochemical conditions in floodplains. En W.J. Junk (eds.), The central Amazon floodplain: ecology of a pulsing system (pp. 69–108). Berlin: Springer Verlag.
Garnier, E., Cortez, J., Billes, G., Navas, M.L., Roumet, C., Debussche, M., Laurent, G., Blanchard, A., Aubry, D., Bellmann, A., Neill, C., y Toussaint, J.P. (2004). Plant functional markers capture ecosystem properties during secondary succession. Ecology, 85, 2630-2637.
Grime, J.P. (1998). Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 86, 902-910.
Higuchi, N., Lima, A.J.N., Teixeira, L.M. (2006). Apostila Biometria Florestal. Manaus. Manaus: Instituto Nacional de Pesquisas da Amazonia.
ICMBIO, 2012. Instituto Chico Mendes de Conservação da Biodiversidade (ICMBIO). Plano de Manejo da Floresta Nacional de Caxiuanã, Volume I. 579p.
Junk, W.J. (2000). Mechanisms for development and maintenance of biodiversity in neotropical floodplains. En B. Gopal, W.J. Junk, y J.A. Davis (eds.), Biodiversity in wetlands: assessment, function and conservation (pp. 119-139). Leiden: Backhuys Publishers.
Junk, W.J., Piedade, M.T.F., Schöngart, J., Wittmann, F. (2012). A classification of major natural habitats of Amazonian whitewater river floodplains (várzeas). Wetlands Ecology and Management, 20, 461–475.
King, D. (1991). Correlations between biomass allocation, relative growth rate and light environment in tropical forest saplings. Functional Ecology, 5, 485–492.
King, D.A., Davies, S.J., y Noor, N.S.M. (2006).Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest. Forest and Ecology Manage, 223, 152–158.
Kurtz, B. C., y Araújo, D. S. D. (2000). Composição florística e estrutura do componente arbóreo de um trecho de Mata Atlântica na Estação Ecológica Estadual do Paraíso, Cachoeiras de Macacu, Rio de Janeiro, Brasil. Rodriguésia, 51, 69-111.
Lenza, E., y Oliveira,P.E. (2006). Biologia reprodutiva e fenologia de Virola sebifera Aubl. (Myristicaceae) em mata mesofítica de Uberlândia, MG, Brasil. Revista Brasileira de Botânica, 29 (3), 443-451.
Malhi, Y., Phillips, O.L., Lloyd, J., Baker, T., Wright, J., Almeida, S., Arroyo, L., Frederiksen, T., Grace, J., Higuchi, N., Killeen, T., Laurance, W.F., Leaño, C., Lewis, S., Meir, P., Monteagudo, A., Neill, D., Vargas, P.N., Panfil, S.N., Patiño, S., Pitman, N., Quesada, C.A., Rudas-Ll., A., Salomão, R., Saleska, S., Silva, N., Silveira, M., Sombroek, W.G., Valencia, R., Martínez, R.V., Vieira, I.C.G., y Vincenti, B. (2002). An international network to monitor the structure, composition and dynamics of Amazonian forests (RAINFOR). Journal of Vegetation Science, 13, 439-450.
Malhi, Y., Baker, T.R., Phillips, O.L., Almeida, S., Alvarez, E., Arroyo, L., Chave, J., Czimczik, C.I., Di Fiori, A., Higuchi, N., Killeen, T.J., Laurance, S.G., Laurance, W.F., Lewis, S.L., Montoya, L.M.M., Monteagudo, A., Neill, D.A., Vargas, P.N., Patiño, S., Pitman, N.C.A., Quesada, C.A., Salomão, R., Silva, J.N.M., Lezama, A.T., Martínez, R.V., Terborgh, J., Vinceti, B., y Lloyd, J. (2004). The above-ground coarse wood productivity of 104 Neotropical forest plots. Global Change Biology, 10, 563-591.
Malhi, Y., Wood, D., Baker, T.R., Wright, J., Phillips, O.L., Cochrane, T., Meir, P., Chave, J., Almeida, S., Arroyo, L., Higuchi, N., Killeen, T.J., Laurance, S.G., Laurance, W.F., Lewis, S.L., Monteagudo, A., Neill, D.A., Vargas, P.N., Pitman, N.C.A., Quesada, C.A., Salomão, R., Silva, J.N.M., Lezama, A.T., Terborgh, J., Martínez, R.V., y Vinceti, B. (2006). The regional variation of aboveground live biomass in old-growth Amazonian forest. Global Change Biology, 12, 1-32.
Melack, J.M., y Hess, L.L. (2010).Remote sensing of the distribution and extent of wetlands in the Amazon basin. En E.J. Junk, M.T.F. Piedade, F. Wittmann, J. Schongart, y P. Parolin (eds.), Amazon floodplain forests: ecophysiology, biodiversity and sustainable management (pp. 43–59). Berlin: Springer Verlag.
Opler, P.A., Bawa, K.S. (1978). Sex ratios in tropical forest trees. Evolution, 32, 812-821.
Phillips, O.L. et al. (1998). Change in the carbon balance of tropical forest: Evidence from long-term plots. Science, 282, 439-442.
Phillips,O.L., Higuchi, N., Vieira, S., Baker, T.R., Chao, K., y Lewis, S.L. (2009). Changes in Amazonian Forest Biomass, Dynamics, and Composition 1980–2002. Amazonia and Global Change: Geophysical Monograph Series, 186, 373-387.
Piccinin, J., y Ruivo, M.L. (2012). Os solos da Floresta Nacional de Caxiuanã. En Instituto Chico Mendes de Conservação da Biodiversidade (eds.), Plano de Manejo da Floresta Nacional de Caxiuanã (pp. 120-127). Brasíli: ICMBIO.
Piña-Rodrigues, F.C.M. (1999). Ecologia reprodutiva e conservação de Virola surinamensis (Rol.) Warb. na região do Estuário Amazônico. (Tese de Doutorado em Ecologia). Universidade Estadual de Campinas, Instituto de Biologia, São Paulo.
Piña-Rodrigues, F.C.M., y Mota, C.G. (2000). Análise da atividade extrativa de virola (Virola surinamensis (Rol.) Warb) no estuário Amazônico. Floresta e Ambiente, 7 (1), 40–5.
Piña-Rodrigues, F.C.M., y Mota, C.G. (1996). Comparação de crescimento entre procedências de Virola surinamensis (Rol) Warb. Floresta Ambient., 3, 49-60.
Salomão, R.P. et al. (2007). Manejo florestal na várzea: caracterização, restrições e oportunidades para sua adoção. En: R.P. Salomão, E.F. Terezo, M.A.G. Jardim (eds), Manejo florestal nas várzeas: oportunidades e desafios (pp. 11-138). Belém: Museu Paraense Emílio Goeldi (Coleção Adolpho Ducke).
Schöngart, J., Piedade, M.T.F., Wittmann, F., Junk, W.J., y Worbes, M. (2005). Wood growth patterns of Macrolobium acaciifolium (Benth.) (Fabaceae) in Amazonian black-water and white-water floodplain forests. Oecologia, 145, 454-461.
Shapiro, S.S., y Wilk, M.B. (1965). An analysis of variance test for normality (complete samples). Biometrika, 52, 591- 611.
Sumida, A., Ito, H., y Isagi, Y. (1997). Trade-off between height growth and stem diameter growth for an evergreen Oak, Quercus glauca, in a mixed hardwood forest. Functional Ecology, 11, 300-309.
Xavier, G.P.S. (2009). Mudanças no estoque de carbono da biomassa lenhosa de florestas de várzea baixa da Amazônia Central ao longo de um gradiente sucessional. (Dissertação de Mestrado em Botânica). Instituto Nacional de Pesquisas Amazônicas. Manaus, Amazonas.
Wolf, L.M. (1997). Differential flower herbivory and gall formation on males and females oí Neea psychotrioicles, a dioecious tree. Biotropica, 29 (2), 169-174.
Worbes, M., (1997). The forest ecosystem of the floodplains. En W.J. Junk (ed), The Central Amazon Floodplains: Ecology of a Pulsing System (pp. 223-266). Berlin: Springer Verlag.
Zar, J.H. (2010). Biostatistical Analysis, 5 ed. New Jersey: Prentice-Hall, Englewood Cliffs.
1. Instituto Federal de Educação, Ciência e Tecnologia do Pará. denise.cunha@ifpa.edu.br
2. Coordenação de Botância. Museu Paraense Emílio Goeldi. lvferreira@museu-goeldi.br