![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 38) Año 2017. Pág. 8
Edilaine DUARTE 1; Ana Carolina da SILVA 2; Pedro HIGUCHI 2; Janaina Gabriela LARSEN 3; Monique Bohora SCHLICKMANN 1; Aline GROSS 1; Luran Monteiro MUZEKA 3; Jaqueline Beatriz Brixner DREYER 1; Guilherme Neto dos SANTOS 3; Silvane de Fátima SIQUEIRA 1; Manoela Bez VEFAGO 1; Felipe Domingos MACHADO 3
Recibido: 17/03/2017 • Aprobado: 23/04/2017
RESUMO: Objetivou-se caracterizar a dinâmica e estrutura de espécies lenhosas regenerantes em uma área degradada altomontana. Foram amostrados indivíduos arbustivo-arbóreos em 2014, 2015 e 2016, sendo construídos histogramas para a distribuição das alturas e, para as espécies de maior densidade, calculados o recrutamento e mortalidade. Ao longo dos anos, o número de indivíduos aumentou e houve variação da distribuição hipsométrica. As cinco espécies mais abundantes apresentaram padrões de dinâmica distintos, sendo que estas apresentam potencial para a restauração de áreas perturbadas. |
ABSTRACT: We aimed to describe the dynamics and structure of woody regenerative tree species in a degraded highland area. Shrub-tree individuals were sampled in 2014, 2015 and 2016. Height distribution histograms were built and recruitment and mortality were determined for the most abundant species. Along time, there were an increment in number of individuals and changes in hypsometric distribution. The five most abundant species, with potential to be used in restoration of degraded lands, showed distinctive dynamics patterns. |

A paisagem natural da região sul do Brasil foi intensamente alterada no passado por práticas de desmatamentos, com a finalidade agropecuária, e pela exploração predatória dos recursos naturais, como das espécies madeireiras. Como consequência, as comunidades vegetais foram impactadas, afetando a manutenção da diversidade biológica existente (Nascimento et al., 2001) e reduzindo o potencial de restauração destas áreas.
Dentre as fitofisionomias que ocorrem nessa região e que foram intensamente exploradas, destaca-se a floresta com araucárias ou Floresta Ombrófila Mista (FOM), considerada uma das mais notáveis em termos de valor ecológico, por abrigar espécies típicas e atributos biológicos únicos em todo o planeta (Oliveira Filho et al., 2015). Devido à grande exploração da araucária (Araucaria angustifolia (Bertol.) Kuntze) e de outras espécies associadas a esta formação florestal (Figueiredo Filho et al., 2011), a FOM, com o passar do tempo, foi gradativamente sendo substituída por culturas agrícolas (Mazon, 2014). Esse cenário resultou na inclusão da fitofisionomia na categoria de criticamente ameaçada e sua espécie típica, a araucária, foi avaliada como em risco de extinção (Medeiros et al., 2005).
De forma geral, devido ao histórico de perturbação, esta formação florestal pode ser considerada, segundo Corlett (1994), como uma floresta secundária. Entretanto, apesar da grande alteração na sua composição florística e estrutural, esta fitofisionomia possui ampla importância ecológica (Silva et al., 2012), atuando de diferentes formas na manutenção da biodiversidade existente.
A FOM é subdividida em quatro subformações (IBGE, 2012), que alteram sua vegetação ao longo de um gradiente altitudinal. Dentre elas, a subformação Alto-Montana, que ocorre em altitudes acima de 1000 m na região (IBGE, 2012), apresenta condições limitantes para o estabelecimento da vegetação, como as baixas temperaturas, os solos pouco férteis e a constante presença de neblina (Higuchi et al., 2013). Como resposta adaptativa a essas condições, espécies que se encontram nestas áreas tendem a apresentar uma vegetação composta por árvores de menor porte, quando comparadas às formações florestais localizadas em altitudes inferiores (Vieira et al., 2014).
Neste sentido, a associação entre a degradação sofrida no passado e os fatores restritivos desta formação, pode ser considerada limitante ao estabelecimento de espécies e à recuperação destas áreas. Assim, Pereira et al. (2001) mencionam que o estudo destas comunidades vegetais é de imprescindível importância, uma vez que contribuem para um maior conhecimento dos processos relacionados à dinâmica, sucessão ecológica e regeneração natural destas áreas perturbadas. Dessa forma, o presente estudo teve por objetivo caracterizar a dinâmica e a estrutura populacional de espécies lenhosas regenerantes em uma área degradada em processo inicial de regeneração no Parque Nacional de São Joaquim, no sul do Brasil. O estudo abrangeu um período de três anos de acompanhamento da regeneração natural, o que pode contribuir na compreensão do processo de formação da comunidade e possibilitou previsões sobre o desenvolvimento futuro da floresta.
O estudo foi conduzido em uma área de campo degradado, em processo inicial de restauração, localizado no Parque Nacional de São Joaquim, entre as coordenadas 28°05'36,42"S e 49°30'17,10"O, no município de Urubici, Santa Catarina, sul do Brasil. A área está situada a uma altitude de 1628 m e apresentava como cobertura original a Floresta Ombrófila Mista Alto-Montana, que foi desmatada na década de 1960 para a formação de pastagem. Em 2007 a área foi desapropriada e protegida, e, atualmente, se encontra em processo inicial de sucessão (regeneração natural pós distúrbio).
O clima da região é classificado como Cfb, segundo Köppen. De acordo com Hijmans et al. (2005), a precipitação e temperatura média anual da região é de 1753 mm e 12,7°C, respectivamente, sendo que as baixas temperaturas do inverno podem promover a ocorrência de geada e neve no local. A vegetação é composta por mosaicos de campos naturais ou antrópicos e a Floresta Ombrófila Mista Alto-Montana, que também é conhecida como floresta nebular devido à presença de neblina constante.
Para o estudo da regeneração natural, foi instalado na área do campo degradado, em 2014, uma transecção, adjacente à um fragmento de Floresta Ombrófila Mista Alto-Montana (Figura 1). Essa transecção possuía 20 m de largura por 100 m de comprimento, e foi subdivida em 20 parcelas permanentes de 10 x 10 m, totalizando 2000 m² de área amostral. Foram amostrados todos os indivíduos arbustivo-arbóreos (lenhosos) com altura ≥ 20 cm, que tiveram suas respectivas alturas medidas por meio de uma régua graduada. Para a análise da dinâmica da comunidade regenerante, foi realizada a re-amostragem dos indivíduos em 2015 e 2016, contabilizando e medindo a altura dos indivíduos sobreviventes, contabilizando os indivíduos mortos e recrutando aqueles que atingiram o nível de inclusão de 20 cm.

Figura 1. Localização da área de estudo no Parque Nacional de São Joaquim, Urubici, sul do Brasil.
Figure 1. Study area location in São Joaquim National Park, municipaity of Urubici, South Brazil
A estrutura da comunidade, para os anos de 2014, 2015 e 2016, foi analisada por meio de histogramas de frequências, com base na distribuição dos indivíduos em classes de altura (nc), pelo método de Sturges: nc = 1+3,33.log(n), em que n = número de indivíduos. O intervalo de classes (L) foi determinado por meio da razão entre a amplitude total e o número de classes: L = R/nc, em que: R = amplitude (maior valor menos o menor valor da amostra).
Para as cinco espécies de maior densidade na área, foram calculadas as taxas de recrutamento (R) e mortalidade (M), por meio das equações descritas por Sheil e May (1996): R = (1-(1-r/Nt)1/t)x100 e M = (1-((No-m)/No)1/t)x100, em que r = número de árvores recrutadas; Nt = número final de árvores depois de t; t = intervalo de tempo entre os inventários; No = número inicial de árvores; m = número de árvores mortas.
No levantamento da regeneração natural foram amostrados, no primeiro inventário (2014), 690 indivíduos, distribuídos em 28 espécies. Em 2015, amostrou-se 841 indivíduos, distribuídos em 29 espécies. No terceiro inventário (2016) foram amostrados 1030 indivíduos, pertencentes a 32 espécies. Esse resultado sugere um incremento mais expressivo no número de indivíduos do que no número de espécies, sugerindo maior estabilidade na riqueza do que na estrutura da comunidade. Esse é um padrão comum em áreas em processo de recuperação inicial, onde há, em um primeiro momento, aumento no número de indivíduos (estruturação da comunidade) e lento incremento no número de espécies. Segundo Machado (2005), isso indica que a comunidade se encontra em processo de estruturação e construção do mosaico silvigenético.
As alturas encontradas para a comunidade variaram de 20 cm a 450 cm, com a maior abundância de indivíduos nas menores classes de altura. Nos dois primeiros anos, 2014 (Figura 2a) e 2015 (Figura 2b), a distribuição hipsométrica tendeu a normal com assimetria positiva. Para o último ano, 2016 (Figura 2c), o padrão observado apresentou uma tendência à distribuição hipsométrica do tipo exponencial negativa, também conhecido como “J invertido”. Enquanto a tendência de distribuição normal dos indivíduos em classes de altura tem sido relatada para florestas em estágio inicial de sucessão (Alves Junior et al., 2007; Costa Junior et al., 2008; Marangon et al., 2008), o padrão em “J invertido” é mais frequente em florestas nativas em avançado estágio sucessional (Gama et al., 2002; Felfili, 2007). Desta forma, fica evidenciado que no processo de sucessão a recuperação da estrutura hipsométrica pode ocorrer de forma rápida. É interessante observar que, em 2016, houve maior número de indivíduos na primeira classe de altura, ao contrário dos demais anos, onde a maior abundância foi observada na segunda classe. Isso ocorreu devido ao aumento de número de indivíduos ao longo dos anos, o que acabou por recrutar mais indivíduos de menor porte.

Figura 2. Estrutura hipsométrica da comunidade lenhosa regenerante de uma área em processo de restauração no Parque Nacional de São Joaquim, Urubici, sul do Brasil, nos três anos de avaliação. Figura 2a (2014): classe 1 = 22,00 a 51,81 cm; classe 2 = 51,82 a 81,63 cm; classe 3 = 81,64 a 111,45 cm; classe 4 = 111,46 a 141,27 cm; classe 5 = 141,28 a 171,09 cm; classe 6 = 171,10 a 200,91 cm; classe 7 = 200,92 a 230,73 cm; classe 8 = 230,74 a 260,55 cm; classe 9 = 260,56 a 290,37 cm; classe 10 = 290,38 a 320,19 cm; classe 11 = 320,20 cm a 350,00 cm; Figura 2b (2015): classe 1 = 20,00 a 53,63 cm; classe 2 = 53,64 a 87,27 cm; classe 3 = 87,28 a 120,91 cm; classe 4 = 120,92 a 154,55 cm; classe 5 = 154,56 a 188,19 cm; classe 6 = 188,20 a 221,83 cm; classe 7 = 221,84 a 255,47 cm; classe 8 = 255,48 a 289,11 cm; classe 9 = 289,12 a 322,75 cm; classe 10 = 322,76 a 356,39 cm; classe 11 = 356,40 a 390,00 cm; Figura 2c (2016): classe 1 = 20,00 a 59,09 cm; classe 2: 59,10 a 98,19 cm; classe 3: 98,20 a 137,29 cm; classe 4 = 137,30 a 176,39 cm; classe 5 = 176,40 a 215,49 cm; classe 6 = 215,50 a 254,59 cm; classe 7 = 254,60 a 293,69 cm; classe 8 = 293,70 a 332,79 cm; classe 9 = 332,80 a 371,89 cm; classe 10 = 371,90 a 410,99 cm; classe 11 = 411,00 a 450,00 cm.
Figure 2. Hypsometric structure of the woody regenerative community in an area under restoration process, at São Joaquim National Park, in the municipality of Urubici, South Brazil, along three years of evaluation. Figure 2a (2014): class 1 = 22,00 to 51,81 cm; class 2 = 51,82 to 81,63 cm; class 3 = 81,64 to 111,45 cm; class 4 = 111,46 to 141,27 cm; class 5 = 141,28 to 171,09 cm; class 6 = 171,10 to 200,91 cm; class 7 = 200,92 to 230,73 cm; class 8 = 230,74 to 260,55 cm; class 9 = 260,56 to 290,37 cm; class 10 = 290,38 to 320,19 cm; class 11 = 320,20 cm to 350,00 cm; Figure 2b (2015): class 1 = 20,00 to 53,63 cm; class 2 = 53,64 to 87,27 cm; class 3 = 87,28 to 120,91 cm; class 4 = 120,92 to 154,55 cm; class 5 = 154,56 to 188,19 cm; class 6 = 188,20 to 221,83 cm; class 7 = 221,84 to 255,47 cm; class 8 = 255,48 to 289,11 cm; class 9 = 289,12 to 322,75 cm; class 10 = 322,76 to 356,39 cm; class 11 = 356,40 to 390,00 cm; Figure 2c (2016): class 1 = 20,00 to 59,09 cm; class 2: 59,10 to 98,19 cm; class 3: 98,20 to 137,29 cm; class 4 = 137,30 to 176,39 cm; class 5 = 176,40 to 215,49 cm; class 6 = 215,50 to 254,59 cm; class 7 = 254,60 to 293,69 cm; class 8 = 293,70 to 332,79 cm; class 9 = 332,80 to 371,89 cm; class 10 = 371,90 to 410,99 cm; class 11 = 411,00 to 450,00 cm.
Do total de indivíduos amostrados no último inventário, 71% (732 indivíduos) pertenceram às cinco espécies mais abundantes: Solanum cassioides L.B.Sm. & Downs (31,94%), Baccharis uncinella DC. (19,22%), Solanum paranense Dusén (8,64%), Myrceugenia regnelliana (O.Berg) D.Legrand & Kausel (5,72%) e Berberis laurina Billb. (5,53%). Espécies da família Solanaceae e Asteraceae, tais como as dos gêneros Baccharis e Solanum, são comuns em áreas com maior altitude em estágio inicial de sucessão (Liebsch et al., 2009). São caracterizadas por serem espécies pioneiras, que apresentam rápido crescimento e frutificação, e que assumem papel importante na colonização imediata após perturbações, promovendo o fechamento de clareiras e facilitando o avanço sucessional. Considerando que Solanum cassiodes, Solanum paranense, Berberis laurina e Myrceugenia regnelliana apresentam dispersão zoocórica (Negrini et al., 2012; Ferreira et al., 2013; Mendes, 2015), infere-se que a fauna desempenhe um papel importante no estabelecimento inicial da vegetação arbórea, o que pode acelerar o processo de restauração. Já Baccharis uncinella apresenta dispersão anemocórica (Negrini et al., 2012), o que facilita sua colonização em áreas abertas.
Considerando o caráter pioneiro das três espécies mais abundantes, a elevada expressividade das mesmas pode ser explicada pelo histórico de perturbação recente da área de estudo. Assim, este resultado indica o elevado potencial de estabelecimento destas espécies após distúrbios em áreas de FOM Alto-Montana, sendo relevantes na sucessão florestal e atuando como facilitadoras no processo de colonização de espécies.
Em relação à representatividade das espécies em cada período analisado, observa-se que a maioria apresentou acréscimo do número de indivíduos entre os anos de avaliação, exceto Solanum paranense e Baccharis uncinella, que tiveram decréscimo da população em 2016 (Figura 3). Além disso, destaca-se que Solanum cassioides apresentou um aumento expressivo em seu tamanho populacional no período 2015-2016.
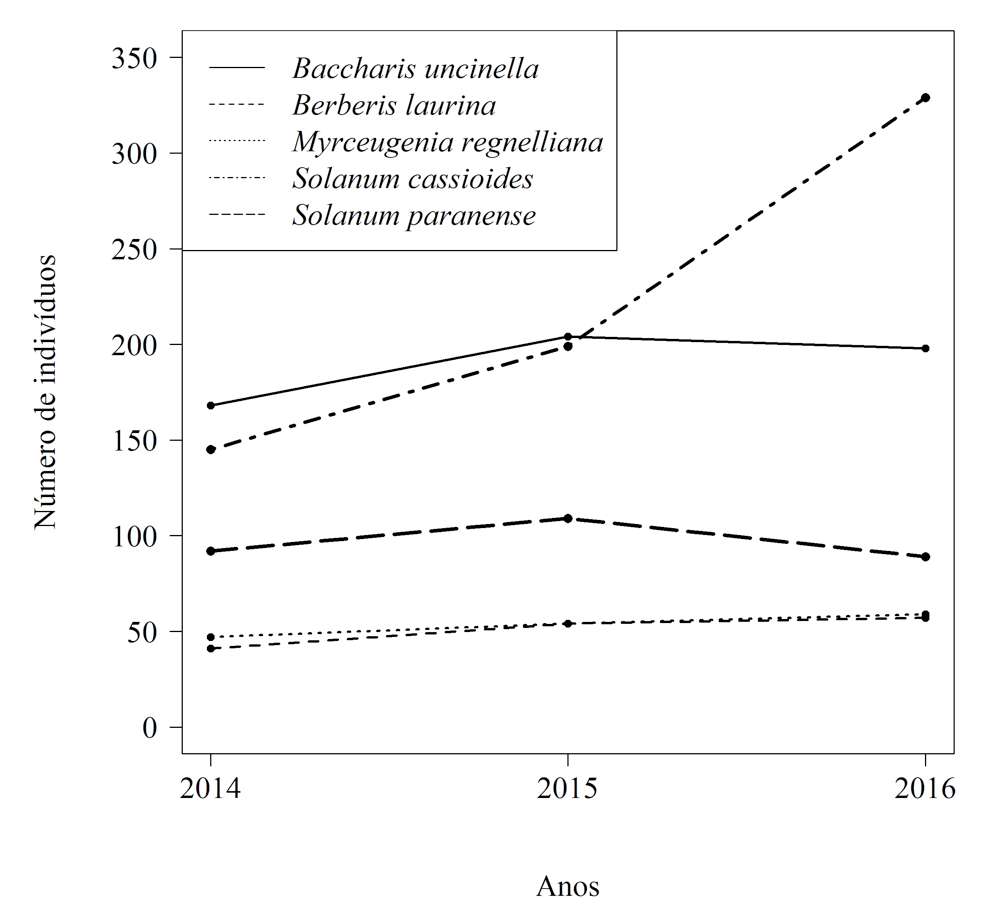
Figura 3. Número de indivíduos das cinco espécies lenhosas de maior abundância em uma área em processo de restauração, durante três anos de avaliação, no Parque Nacional de São Joaquim, Urubici, sul do Brasil.
Figure 3. Number of individuals of the five most abundant woody species in an area under restoration process, along three years of evaluation, at São Joaquim National Park, in the municipality of Urubici, South Brazil.
Este padrão é confirmado quando se analisa as taxas de mortalidade e recrutamento das espécies (Tabela 1). Apesar de Baccharis uncinella e Solanum paranense, que são espécies pioneiras,apresentarem, no período 2014-2015, maiores taxas de recrutamento do que de mortalidade, seguindo o mesmo padrão das demais espécies avaliadas, no período 2015-2016 o padrão se inverte para essas duas espécies, que apresentaram maiores valores de mortalidade. Solanum cassioides, também uma espécie pioneira, obteve, para o mesmo período, maior recrutamento do que mortalidade, sendo que esta apresentou valor bastante superior de recrutamento no último período (62,92%.ano-1).
Tabela 1. Taxas de mortalidade e recrutamento das cinco espécies lenhosas de maior abundância em uma área em processo de restauração no Parque Nacional de São Joaquim, Urubici, sul do Brasil. Em que: M = Taxa de mortalidade (%.ano-1); R = Taxa de recrutamento (%.ano-1).
Table 1. Mortality and recruitment rates of the five most abundant woody species in an area under restoration process at São Joaquim National Park, in the municipality of Urubici, South Brazil. M = mortality rate (%.year-1); R = recruitment rate ((%.year-1).
Períodos |
2014-2015 |
2015-2016 |
||
Espécies |
M |
R |
M |
R |
Baccharis uncinella |
1,79 |
19,51 |
11,71 |
8,59 |
Berberis laurina |
2,44 |
27,27 |
3,64 |
7,02 |
Myrceugenia regnelliana |
0,00 |
12,96 |
1,85 |
10,17 |
Solanum cassioides |
13,10 |
37,31 |
39,30 |
62,92 |
Solanum paranense |
14,13 |
27,52 |
29,36 |
13,48 |
Da mesma forma que observado por Van Breugel et al. (2007), os resultados encontrados evidenciam que, mesmo em sua fase inicial e em um curto período de tempo (três anos), o processo de sucessão pode ser caracterizado por flutuações no tamanho populacional das espécies e, consequentemente, mudanças na representatividade das mesmas. Estas mudanças ocorrem em reposta a uma complexa relação entre fatores bióticos (e.g., banco de sementes, fonte de propágulos, predação de sementes) e abióticos (e.g., ambiente lumínico, propriedades do solo) (Guariguata & Ostertag, 2001), de forma que espécies que se estabelecem incialmente podem exercer a função de facilitadoras para outras mais tardias.
O processo de restauração florestal inicial foi caracterizado por uma rápida recuperação estrutural (aumento na abundância e alteração na distribuição dos indivíduos em clases de altura), porém, com estabilidade na riqueza de espécies. As cinco espécies de maior abundância na comunidade apresentaram padrões de dinâmica distintos, o que pode estar associado a flutuações populacionais, comuns no processo de sucessão florestal. Infere-se que as espécies Solanum cassioides, Baccharis uncinella, Solanum paranense, Myrceugenia regnelliana e Berberis laurina, por apresentarem elevada representatividade no componente regenerativo em uma área de Floresta Ombrófila Mista Alto-Montana em fase inicial de sucessão, possuem grande potencial para serem empregadas em planos de restauração florestal.
À FAPESC (Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina), pela concessão de bolsa à primeira autora do trabalho. Ao CNPq, pelo financiamento do projeto e pela concessão de bolsa de produtividade para o segundo e o terceiro autores. Ao Instituto Chico Mendes de Conservação da Biodiversidade, à equipe do PARNA São Joaquim e ao MSc. Michel Omena, pela permissão para execução do trabalho no Parque Nacional de São Joaquim.
Alves Junior, F. T., Brandão, C. F. L., Rocha, K. D. D., Silva, J. T. D., Marangon, L. C., & Ferreira, R. L. C. (2007). Estrutura diamétrica e hipsométrica do componente arbóreo de um fragmento de Mata Atlântica, Recife-PE. Cerne, 13(1), 83-95.
Corlett, R. T. (1994). What is secondary Forest? Journal of Tropical Ecology, 10(3), 445-447.
Costa Junior, R. F., Ferreira, R. L. C., Rodal, M. J. N., Feliciano, A. L. P., Marangon, L. C., & Silva, W. C. (2008). Estrutura fitossociológica do componente arbóreo de um fragmento de Floresta Ombrófila Densa na mata sul de Pernambuco, nordeste do Brasil. Ciência Florestal, 18(2), 173-183.
Ferreira, P. I., Gomes, J. P., Batista, F., Bernardi, A. P., Bortoluzzi, R. L. C., & Mantovani, A. (2013). Espécies potenciais para recuperação de Áreas de Preservação Permanente no Planalto Catarinense. Revista Floresta e Ambiente, 20(2), 173-182.
Felfili, J. M. (1997). Diameter and height distributions in a gallery forest tree community and some of its main species in central Brazil over a six-year period (1985-1991). Brazilian Journal of Botany, 20(2), 155-162.
Figueiredo Filho, A., Orellana, E., Nascimento, F., Dias, A. N., & Inoue, M. T. (2011). Produção de sementes de Araucaria angustifolia em plantio e em floresta natural no Centro-Sul do Estado do Paraná. Floresta, 41(1), 155-162.
Gama, J. R. V., Botelho, S. A., & Bentes-Gama, M. D. M. (2002). Composição florística e estrutura da regeneração natural de floresta secundária de várzea baixa no estuário amazônico. Revista Árvore, 26(5), 559-566.
Guariguata, M. R., & Ostertag, R. (2001). Neotropical secondary forest succession: changes in structural and functional characteristics. Forest Ecology and Management, 148(1-3), 185-206.
Higuchi, P., Silva, A. C., Almeida, J. A., Bortoluzzi, R. L. C., Mantovani, A., Ferreira, T. S., Souza, S. T., Gomes, J. P., & Silva, K. M. (2013). Florística e estrutura do componente arbóreo e análise ambiental de um fragmento de Floresta Ombrófila Mista Alto-Montana no município de Painel, SC. Ciência Florestal, 23(1), 153-164.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., & Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25(15), 1965-1978.
IBGE. (2012). Manual técnico da vegetação brasileira. Rio de Janeiro: Fundação Instituto Brasileiro de Geografia e Estatística.
Liebsch, D., Mikich, S. B., Possete, R. S., & Ribas, O. S. (2009). Levantamento florístico e síndromes de dispersão em remanescentes de Floresta Ombrófila Mista na região centro-sul do estado do Paraná. Hoehnea, 36(2), 233-248.
Machado, E. L. M. (2005). Heterogeneidade espacial e temporal em um fragmento de floresta estacional em Lavras, MG. (Dissertação de Mestrado). Universidade Federal de Lavras. Lavras.
Marangon, L. C., Feliciano, A. L. P., Brandão, C. F. L. S., & Alves Júnior, F. T. (2008). Relações florísticas, estrutura diamétrica e hipsométrica de um fragmento de Floresta Estacional Semidecidual em Viçosa (MG). Floresta, 38(4), 699-709.
Mazon, J. A. (2014). Composição florística e fitossociológica de Floresta Ombrófila Mista em áreas sob manejo silvipastoril e sucessão secundária. (Dissertação de Mestrado). Universidade Estadual do Centro Oeste. Guarapuava.
Medeiros, J. D., Savi, M., & Brito, B. F. A. (2005). Seleção de áreas para criação de Unidades de Conservação na Floresta Ombrófila Mista. Biotemas, 18(2), 33-50.
Mendes, A. R. (2015). Floresta ciliar do rio pelotas ao longo de um gradiente altitudinal: fitossocilogia e variáveis edáficas. (Dissertação de Mestrado). Universidade do Estado de Santa Catarina, Lages.
Nascimento, A. R. T., Longhi, S. J., & Brena, D. A. (2001). Estrutura e padrões de distribuição espacial de espécies arbóreas em uma amostra de Floresta Ombrófila Mista em Nova Prata, RS. Ciência Florestal, 11(1), 105-119.
Negrini, M., Aguiar, M. D., Vieira, C. T., Silva, A. C., & Higuchi, P. (2012). Dispersão, distribuição espacial e estratificação vertical da comunidade arbórea em um fragmento florestal no Planalto Catarinense. Revista Árvore, 36(5), 919-929.
Oliveira Filho, A. T., Budke, J. C., Jarenkow, J. A., Eisenlohr, P. V., & Neves, D. R. M. (2015). Delving into the variations in tree species composition and richness across South American subtropical Atlantic and Pampean forests. Journal of Plant Ecology, 8(3), 242-260.
Pereira, I. M., Andrade, L. A, Costa, J. R. M, & Dias, J. M. (2001). Regeneração natural em um remanescente de Caatinga sob diferentes níveis de perturbação, no Agreste Paraibano. Acta Botânica Brasílica, 15(3), 413-426.
Sheil, D., & May, R. M. (1996). Mortality and recruitment rate evaluations in heterogeneous tropical forests. Journal of Ecology, 84(1), 91-100.
Silva, C., Higuchi, P., Aguiar, M. D., Negrini, M., Neto, J. F., & Hess, A. F. (2012). Relações florísticas e fitossociologia de uma Floresta Ombrófila Mista Montana secundária em Lages, Santa Catarina. Ciência Florestal, 22(1), 193-206.
Van Breugel, M., Bongers, F., & Martínez‐Ramos, M. (2007). Species dynamics during early secondary forest succession: recruitment, mortality and species turnover. Biotropica, 39(5), 610-619.
Viera, R. S., Blum, C. T., & Roderjan, C. V. (2014). Caracterização florística e estrutural de uma Floresta Ombrófila Densa Alto-Montana na Serra do Capivari, Campina Grande do Sul, Paraná. Floresta, 44(4), 565-576.
1. Mestrandos em Engenharia Florestal, na linha de pesquisa Ecologia de espécies florestais e ecossistemas associados. Departamento de Engenharia Florestal. Universidade do Estado de Santa Catarina. E-emails de contato: ediduarte10@gmail.com, nique_bn@hotmail.com , alinegros@hotmail.com , jaqueline.bbdreyer@gmail.com , biosilvane@hotmail.com , manuvefago@gmail.com
2. Doutores em Engenharia Florestal, atualmente professores da área de Dendrologia, Ecologia e Fitossociologia. Departamento de Engenharia Florestal. Universidade do Estado de Santa Catarina. E-emails de contato: carol_sil4@yahoo.com.br , higuchip@gmail.com
3. Graduandos em Engenharia Florestal, atuando na área de Ecologia Florestal e Dendrologia. Departamento de Engenharia Florestal. Universidade do Estado de Santa Catarina. E-emails de contato: janagabrielalarsen@hotmail.com , luranmuzeka@gmail.com , guiguis_netto@hotmail.com , felipe.d.machado@hotmail.com