![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 36) Año 2017. Pág. 29
Igor DO VALE 1; Izildinha Souza MIRANDA 2; Danielle MITJA 3; Graciliano Galdino Alves dos SANTOS 4; Alessio Moreira dos SANTOS 5; Tâmara Thaiz Santana LIMA 6 Luiz Gonzaga Silva COSTA 7
Recibido: 01/03/2017 • Aprobado: 20/03/2017
RESUMO: A variação da riqueza de espécies entre diferentes mosaicos rurais e tipos de uso da terra foi analisada em escala local e da paisagem. Nove tipos de uso entre três mosaicos rurais foram comparados através de curvas de rarefação estabelecidas para três estratos verticais da vegetação. A riqueza foi fortemente influenciada pela composição dos tipos de uso implantados em cada fazenda na paisagem. Florestas secundárias tiveram maior riqueza em mosaicos com maior cobertura florestal de alta riqueza na paisagem. |
ABSTRACT: The variation of species richness between different rural mosaics and land-use types was analyzed on a local and landscape scale. Nine land-use types among three rural mosaics were compared through rarefaction curves established in three vertical strata of the vegetation. Richness was strongly influenced by the composition of the land-use types implanted in each farm within the landscape. Secondary forests had greater richness in mosaics with higher high richness forest cover in the landscape. |

Em função da conversão de florestas para áreas de cultivos agrícolas e pastagens na Amazônia, mosaicos de diferentes coberturas vegetais tem surgido, principalmente em áreas de agricultura familiar. A heterogeneidade encontrada nesses mosaicos, onde fragmentos de diferentes cultivos são conectados com áreas naturais, supre uma diversidade de necessidades biológicas e ecológicas (como refúgio, alimentação, e corredores de dispersão) que auxiliam na sobrevivência e diversidade das espécies, seja medida em grande ou pequena escala (Benton et al., 2003).
Estudos que focam no efeito da modificação e remoção das coberturas tropicais sobre a biodiversidade têm aumentado nesses mosaicos (Gibson et al., 2011; Newbold et al., 2015; De Castro Solar et al., 2016); pois quantificar e compreender as principais respostas da biodiversidade às perturbações humanas em escalas múltiplas é fundamental para promover planos de conservação (De Castro Solar et al., 2016).
Em geral, essas mudanças nos usos da terra reduzem a biodiversidade, embora o tamanho do efeito varia de acordo com a região, grupo taxonômico, métricas ecológicas e tipo de perturbação, além da escala de estudo (Gibson et al., 2011; Chisholm et al., 2013).
A diversidade alfa, derivada da riqueza de espécies em um determinado local, e a diversidade beta, derivada das diferenças na composição de espécies de um local para outro (Crist et al., 2003), podem revelar padrões espaço-temporais contrastantes e a extensão em que a diversidade alfa pode predizer a diversidade beta pode diferir, pois dois locais com igual riqueza de espécies podem compartilhar todas ou nenhuma de suas espécies (Tylianakise et al., 2005; Kessler et al., 2009). Assim, comparar a diversidade alfa e beta em escala local e de paisagem é uma área importante e ainda pouco compreendida da pesquisa ecológica básica e aplicada (Kessler et al., 2009).
O desenvolvimento de planos de conservação eficazes para paisagens multifuncionais exige uma compreensão do valor das florestas secundárias transitórias na manutenção da diversidade vegetal (Gardner et al., 2007; Melo et al., 2013; Van Breugel et al., 2013). As práticas de uso da terra podem mudar fortemente as trajetórias de recuperação dessas florestas (Mesquita et al., 2015; Arroyo-Rodriguez et al., 2015) e, a longo prazo, a intensificação do uso da terra pode reduzir sua resiliência, o que diminui o potencial de prestação de serviços ecossistêmicos (Jakovac et al., 2015). Em geral, tipos de uso agrícolas (abandonadas e ativas) têm um impacto muito maior do que os sistemas agroflorestais e plantações, com e sem sombreamento (Gibson et al., 2011).
Uma forma de acessar o efeito das mudanças da cobertura florestal na biodiversidade é a comparação da riqueza entre os diferentes tipos de uso da terra, que pode servir como uma métrica simples e eficaz, para avaliar o valor de conservação de cada tipo de uso da terra e os impactos relativos a diferentes padrões de modificação na paisagem (Mendes et al., 2008; Gibson et al., 2011). Em estudos com plantas, a importância relativa dos diferentes gradientes ambientais e sua interação na determinação da riqueza de espécies de diferentes formas de crescimento precisa ser mais aprofundada (Pausas e Austin, 2001).
As condições e recursos disponíveis em diferentes estratos estabelecem nichos diferenciados para as plantas (Puig, 2008). A partir do estádio juvenil até o adulto, os indivíduos de uma população vegetal passam por uma série de filtros ambientais, que resultam em comunidades de estrutura e composição distintas (Puig, 2008; Carvalho et al., 2009). Além disso, os estratos inferiores, que representam a dinâmica de regeneração, são definidos pelo regime de perturbação (incluindo a intensidade, freqüência e escala de perturbação) e pela biologia das espécies (Kennard et al., 2002). Enquanto, os estratos superiores, presentes nos fragmentos florestais, constituem importante fonte de própagulos e sementes florestais (Vieira e Proctor, 2007). Desta forma, o estudo de diferentes estratos pode auxiliar na compreensão do potencial de conservação biológica dos diferentes tipos de uso do solo.
Neste estudo, objetivamos comparar a diversidade de plantas em diferentes tipos de uso da terra estabelecidos em três mosaicos rurais do sudeste do Estado do Pará, com a finalidade de responder três perguntas: (1) qual a importância da escala (alfa e beta) sobre a diversidade total dos mosaicos? (2) como a riqueza de espécies responde aos diferentes tipos de uso? (3) os padrões de riqueza serão o mesmo em diferentes estratos?
O estudo foi realizado em 2008, em três comunidades rurais com pequenas propriedades familiares: Palmares II, Maçaranduba e Travessão 338-S. Cada comunidade foi considerada como um mosaico rural, onde fragmentos de áreas naturais estão conectados a diferentes tipos de usos da terra. As comunidades estão localizadas no sul e sudeste do Estado do Pará, na região conhecida como Arco do Desmatamento. A perda de biodiversidade nessa região é intensa, e impulsonada pelas mudanças de uso da terra causada pelo desmatamento para a agricultura. Além disso, os incêndios antropogênicos e os graves eventos de seca da última década nesta região também estão impulsionando mudanças florísticas e estruturais e contribuindo para o próprio desmatamento (Morandi et al., 2016).
O Projeto de Assentamento Palmares II (625,263.4 9,343,195.9; UTM, WGS84), localizado no município de Parauapebas, foi criado em 1996, com fazendas que possuem em média 26 ha de área. A comunidade de Maçaranduba (679,602.5 9,473,375.9 UTM, WGS84) está localizada no Projeto de Assentamento Agroextrativista Praia Alta-Piranheiras, no município de Nova Ipixuna. Esta comunidade foi criada em 1994, e suas fazendas tem área média de 71 ha. A comunidade do Travessão 338-S (493,585.7 9,582,189.4 UTM, WGS84) está localizada ao longo da Rodovia Transamazônica, no município de Pacajá. Foi criado em 2001, as fazendas têm tamanho médio de 86 ha.
Essas três comunidades rurais já foram estudadas por De Sartre (2011), Oszwald et al. (2011; 2012), Costa et al. (2012), Grimaldi et al. (2014), Marichal et al. (2014), Do Vale et al. (2015) e Lavelle et al. (2016). A vegetação primária dominante na região é floresta tropical úmida de terra firme, e a temperatura média anual na área é de 26 °C.
As fazendas estudadas são marcadas por uma dinâmica da paisagem recente, apresentando mosaico de diferentes coberturas, tais como pastagens, roças, florestas secundárias, entre outros. No entanto, as três áreas apresentaram dinâmicas de colonização diferenciadas. Uma análise das métricas das paisagens de 2007, e sua dinâmica entre 1990 e 2007, foi apresentada por Oszwald et al. (2011), que inclui um mapa de localização das fazendas e imagens Landsat da cobertura vegetal em 2007. A comunidade de Palmares II apresentava uma paisagem agrícola muito fragmentada, com muitos pastos e roças, além da presença de queimadas recentes nas áreas florestais; Palmares II possui grande rede viária, no entanto, a distância entre o local de residência (Vila de Palmares II) e o local dos cultivos é um fator importante que interfere no grau de impacto humano.
A comunidade de Maçaranduba é mais antiga e apresentava uma paisagem predominantemente agrícola, composta por pastagens em diferentes estágios de desenvolvimento; as florestas secundarias foram formadas após o abandono das áreas agrícolas e as florestas pristinas são remanescentes de matas ciliares ou em áreas de difícil acesso. Maçaranduba é caracterizada por uma passagem muito rápida e intensa da fase florestal à fase agrícola, principalmente pastagens. A comunidade do Travessão 338-S é a mais recentemente afetada pelo desmatamento, as primeiras intervenções agrícolas começaram no início de 1990 e a conversão de florestas em terras agrícolas permanecia ainda marginal em 2007; as fazendas eram principalmente florestais ou compostas por manchas florestais de tamanho grande, o que caracterizava a paisagem do mosaico como florestal (Oszwald et al., 2011).
Uma pesquisa socioeconômica realizada com 50 pequenos agricultores familiares ajudou a definir quais fazendas eram mais representativas dos mosaicos (De Sartre, 2011); e, as nove fazendas mais representativas em cada mosaico foram selecionadas para este estudo.
Em cada fazenda foram estabelecidos cinco pontos amostrais para o inventário da vegetação. Os pontos foram igualmente espaçados ao longo da maior diagonal da fazenda, com o auxílio de um mapa. Assim, a distância entre os pontos foi igual a 1/6 do comprimento da diagonal e variou conforme a fazenda (entre 200 e 400 m). Como os pontos foram espaçados ao longo da diagonal de cada fazenda, eles não foram selecionados com base no tipo de uso da terra. Assim, foi assumido que a cobertura vegetal dos pontos refletiu os tipos de uso da terra predominantes em cada mosaico rural estudado, o que resultou em um número heterogêneo de pontos amostrais para cada tipo de uso por mosaico (Tabela 1).
Foram identificados nove tipos de uso da terra nos três mosaicos rurais estudados. O mosaico de Palmares II, é o único que possui florestas queimadas e não possui florestas conservadas, e tem o maior número de cultivos anuais. Maçaranduba possui o maior número de pastos, enquanto Travessão 338-S tem o maior número de florestas e o único que possui plantios arbóreos. É importante estabelecer que todos os cultivos agrícolas estavam ativos na época da coleta dos dados, que ocorreu em 2008 (Tabela 1).
Em cada ponto amostral a vegetação foi coletada em três estratos. Uma parcela de 10 x 50 m (500 m²) foi alocada para o inventário do estrato superior (indivíduos com DAP > 10 cm), uma subparcela de 5 x 50 m para o estrato médio (indivíduos com DAP < 10 cm e altura > 2,0 m) e finalmente, no centro da parcela, foram distribuídas regularmente 10 subparcelas de 1 x 1 m para o inventário do estrato inferior (indivíduos com 2,0 m < altura > 10 cm). O número de amostras por estrato variou por mosaico, visto que existem tipos de uso que não possuem todos os estratos.
A pré-identificação das espécies foi realizada por um parataxônomo e confirmada por comparação no Herbário João Murça Pires do Museu Paraense Emilio Goeldi. As plantas foram classificadas de acordo com o sistema Angiosperm Phylogeny Group III. Os poucos indivíduos não identificados (2, 5 e 6% dos indivíduos em Palmares II, Maçaranduba e Travessão 338-S, respectivamente) foram excluídos das análises.
Tabela 1. Número de pontos amostrais por tipo de uso em três mosaicos rurais
(PII = Palmares II, M = Maçaranduba, T338S = Travessão 338-S)
na região do “arco do desmatamento”, Amazônia Oriental.
Sigla |
Tipo de Uso da Terra |
Descrição |
Mosaicos Rurais |
Total |
||
PII |
M |
T338 |
|
|||
FloCon |
Florestas Conservadas |
Florestas com estrutura vertical bem definidas e sem sinais de distúrbio humano |
- |
8 |
7 |
15 |
FloExp |
Florestas Alteradas |
Florestas que possuem sinais de perturbação, como clareiras resultantes de retirada de madeira |
5 |
3 |
16 |
24 |
FLoQue |
Florestas Queimadas |
Florestas com sinais do efeito do fogo |
10 |
- |
- |
10 |
SecAnt |
Florestas Secundárias Antigas |
Florestas secundárias com estrato superior arbóreo bem definido, e idade acima de 12 anos |
3 |
9 |
4 |
16 |
SecJov |
Florestas Secundárias Jovens |
Florestas secundárias sem estrato superior arbóreo definido e, idade abaixo de 12 anos |
6 |
4 |
5 |
15 |
PasInv |
Pastos Invadidosa |
Pastos com alta abundância de indivíduos herbáceos e jovens plântulas de espécies lenhosas |
4 |
11 |
6 |
21 |
PasLim |
Pastos Limposa |
Pastos com baixa densidade de espécies lenhosas |
7 |
9 |
1 |
17 |
CulAnu |
Cultivos Anuais |
Arroz (Oryza sp.), feijão (Vigna sp.), milho (Zea mays) e especialmente plantações de mandioca (Manihot esculenta) |
10 |
1 |
2 |
13 |
PlaArb |
Plantio arbóreob |
Plantações de cacau (Theobroma cacao), geralmente combinado com paricá (Schizolobium amazonicum) (dois estratos verticais) |
- |
- |
4 |
4 |
|
Total |
Total de pontos amostrais |
45 |
45 |
45 |
135 |
a Urochloa brizantha foi a forrageira dominante em Maçaranduba e Palmares II, enquanto Urochloa decumbens predominou em Travessão 338-S.
b Plantações arbóreas são comuns no sudeste do Pará e foram restritas ao mosaico de Travessão 338-S, parte integrante da região produtora de cacau do sudeste do Pará.
Riqueza de espécies foi a medida de diversidade utilizada neste estudo, pois além de ser a medida mais utilizada possui algumas vantagens: 1) captura a essência da biodiversidade porque riqueza se associa prontamente à presença de uma variedade de espécies, 2) seu significado é amplamente compreendido, 3) é um parâmetro mensurável, e 4) muitos dados sobre riqueza já existem (Mendes et al., 2008).
Assim, para verificar a importância da escala (alfa e beta) sobre a diversidade total dos mosaicos rurais, foi utilizado a partição aditiva da diversidade (Lande, 1996), onde ɣ = α + β1 + β2, em que a diversidade alfa (α) considera a riqueza total por ponto amostral, diversidade beta considera a diversidade entre pontos amostrais da mesma fazenda (β1) e entre as fazendas amostradas (β2), e diversidade gama (ɣ) é a soma de todas as diversidades anteriores e representa a diversidade do mosaico. Esta abordagem permite que a partição aditiva da diversidade total numa região seja desagregada em componentes específicos da escala da diversidade, que podem ser comparados diretamente; além disso, pode ajudar a entender quais fatores estão controlando a distribuição espacial da biodiversidade (Veech et al., 2002). A partição aditiva foi realizada na interface R-Studio para R (R Foundation for Statistical Computing, Vienna, AT) utilizando a função de adipart do pacote vegan.
Para comparar a riqueza entre os diferentes mosaicos e entre tipos de uso da terra dentro de um mesmo mosaico, a riqueza foi estimada pelo método de rarefação para cada estrato inventariado, utilizando o estimador de Mao Tao, baseado no número de amostras (Colwell, 2013). As curvas de rarefação foram utilizadas, pois o número de amostras entre os tipos de usos e entre os mosaicos foram diferentes; e, também por esse motivo, a riqueza estimada foi extrapolada de forma a homogeneizar o número de amostras e facilitar a comparação. A extrapolação foi realizada segundo Colwell et al. (2012) e tomou como base o tipo de uso ou o estrato com o maior número de amostras.
Tipos de uso com menos de três pontos amostrais não foram considerados na análise. As estimativas e a extrapolação de riqueza foram realizadas no programa Estimates 9.1 (Colwell, 2013). A significância das diferenças observadas na riqueza estimada foi avaliada comparando visualmente as curvas de rarefação por amostras e seus respectivos intervalos de confiança a 95% plotadas em um eixo x de abundância de indivíduos, conforme recomendado por Gotelli e Cowell (2001). No caso de sobreposição dos intervalos de confiança então as riquezas foram consideradas similares, caso contrário foram consideradas diferentes.
A diversidade beta entre as fazendas contribuiu mais fortemente para a riqueza total de espécies dos mosaicos (ou diversidade gama) e dos tipos de uso. Nos mosaicos, a diversidade alfa teve menor contribuição que nos tipos de uso. A contribuição da diversidade beta entre os pontos amostrais foi maior nos mosaicos que entre os tipos de uso (Figura 1).
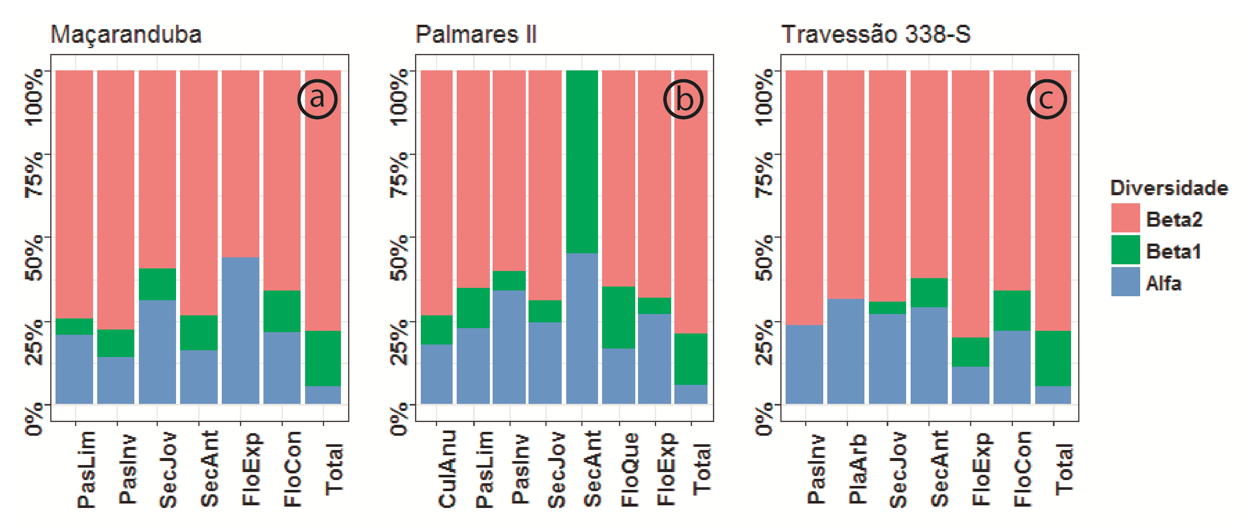
Figura 1. Partição aditiva da diversidade florística (em %) em diferentes tipos de uso da terra em três mosaicos rurais (a = Palmares II, b = Maçaranduba, c = Travessão338-S) na região do “arco do desmatamento”, Amazônia Oriental. Azul = diversidade α (riqueza total por ponto amostral); verde = diversidade β1 (riqueza entre pontos) e rosa = diversidade β2 (riqueza entre fazendas). Siglas para tipos de uso como na Tabela 1.
As florestas secundárias antigas de Palmares II não apresentaram diversidade beta entre fazendas, porque foram presentes em apenas uma fazenda. As florestas exploradas de Maçaranduba, os pastos invadidos e plantios arbóreos do Travessão 338-S não apresentaram diversidade entre pontos amostrais, porque esses tipos de uso apresentaram apenas um ponto amostral em cada fazenda (Figura 1).
Dois fatores podem ter contribuído para o aumento da diversidade beta: a distância entre as fazendas e a fragmentação da paisagem causada pela implantação de diferentes tipos de uso da terra em cada fazenda. Locais próximos uns dos outros são normalmente mais semelhantes em condições ambientais do que locais distantes. Quanto maior for o grau de heterogeneidade espacial subjacente na paisagem, maior será o grau de dissimilaridade da comunidade entre os fragmentos de habitat (tipos de uso da terra) separados por distâncias crescentes (Fortin e Dale, 2005; Sfair et al., 2016). As diferentes condições ambientais entre as fazendas são acentuadas, não só pela distância, mas também pelas escolhas de manejo aplicadas por cada pequeno agricultor familiar.
O tamanho, o local e o tipo de uso das manchas agrícolas em cada fazenda, é uma escolha do produtor, e vai determinar a composição e a estrutura da paisagem. E, quando a mesma quantidade de habitat é distribuída em uma paisagem, através da fragmentação, em vez de permanecer como um grande fragmento único, a diversidade beta é aumentada (Tscharntke et al., 2012). Portanto, a dissimilaridade das comunidades locais (diferentes tipos de uso em diferentes fazendas) determina a biodiversidade em toda a paisagem e substitui os efeitos locais negativos da fragmentação do habitat na biodiversidade – hipótese da dominância da diversidade beta (Tscharntke et al., 2012; Arroyo-Rodriguez et al., 2015).
Entretanto, ao comparar comunidades de abelhas e vespas ao longo de um gradiente de uso da terra, Tylianakis et al. (2005) descobriram que embora a diversidade alfa fosse maior nos agroecossistemas intensivamente utilizados, a diversidade beta foi significativamente maior em sistemas de menor intensidade. Apesar das diferenças de intensidade de uso entre os mosaicos rurais e os tipos de uso da terra, nenhum padrão foi encontrado para as plantas analisadas neste estudo.
Houveram diferenças significativas nas curvas de rarefação de espécies entre os tipos de uso de cada mosaico; contudo, essas diferenças ocorreram apenas nos estratos inferior e médio (Figura 2; Tabela 2). No geral, as florestas conservadas, exploradas e queimadas apresentaram o maior número de espécies entre os tipos de usos de todos os mosaicos, enquanto os pastos invadidos, limpos e cultivos anuais apresentaram a menor riqueza (Figura 2; Tabela 2).
Como sabemos, o maior número de espécies nas florestas é devido à um enriquecimento gradual de espécies durante a sua formação, acompanhado de um aumento em complexidade estrutural e funcional, o que inclui maior estratificação (Puig, 2008; Chazdon 2012).
As florestas secundárias antigas de Maçaranduba e Travessão 338-S, também mostraram a mesma complexidade, pois apresentaram riqueza similar às florestas conservadas e exploradas em todos os estratos (Figura 2; Tabela 2). No estrato inferior de Maçaranduba, apresentaram até mais espécies que as florestas conservadas (Figura 2h; Tabela 2).
Já em Palmares II, as florestas secundárias antigas tiveram riqueza semelhante às florestas queimadas e exploradas apenas no estrato superior, mas foi neste estrato que as florestas queimadas e as capoeiras antigas mostraram baixo número de indivíduos quando comparadas às florestas exploradas (Figura 2c; Tabela 2). Nos estratos inferior e médio, as florestas secundárias antigas foram semelhantes às florestas secundárias jovens, pastos invadidos e cultivos anuais (Figura 2a-b; Tabela 2).
Tabela 2. Riqueza dos estratos inferior, médio e superior dos tipos de uso da terra (ver siglas na Tabela 1) de três mosaicos rurais (Palmares II, Maçaranduba, Travessão338-S) na região do “arco do desmatamento”, Amazônia Oriental. N = Número de amostras, Ind = número de indivíduos, S = espécies inventariadas, S-est = número estimado de espécies, Sd = desvio padrão estimado.
Tipo de uso |
Estrato Inferior |
Estrato Médio |
Estrato Superior |
||||||||||||
n |
Ind |
S |
S-est |
SD |
n |
Ind |
S |
S-est |
SD |
n |
Ind |
S |
S-est |
SD |
|
Palmares II |
|||||||||||||||
FloExp |
5 |
647 |
151 |
245 |
18 |
5 |
866 |
118 |
180 |
14 |
5 |
93 |
51 |
81 |
10 |
FloQue |
10 |
1024 |
239 |
239 |
12 |
10 |
2416 |
150 |
150 |
9 |
10 |
90 |
45 |
45 |
5 |
SecAnt |
3 |
441 |
69 |
120 |
15 |
3 |
478 |
43 |
69 |
11 |
3 |
32 |
14 |
28 |
8 |
SecJov |
6 |
2157 |
119 |
168 |
13 |
6 |
1462 |
70 |
91 |
8 |
6 |
1 |
1 |
1 |
0 |
PasInv |
4 |
1224 |
99 |
155 |
14 |
4 |
354 |
24 |
46 |
9 |
4 |
5 |
4 |
7 |
3 |
PasLim |
7 |
1417 |
73 |
92 |
9 |
7 |
220 |
12 |
15 |
4 |
10 |
3 |
2 |
2 |
1 |
CulAnu |
10 |
3161 |
145 |
145 |
9 |
10 |
474 |
32 |
32 |
4 |
- |
- |
- |
- |
- |
Maçaranduba |
|||||||||||||||
FloCon |
8 |
1151 |
143 |
171 |
10 |
8 |
644 |
161 |
196 |
12 |
8 |
168 |
64 |
69 |
6 |
FloExp |
3 |
425 |
85 |
190 |
25 |
3 |
319 |
102 |
189 |
22 |
3 |
93 |
32 |
52 |
9 |
SecAnt |
9 |
1024 |
185 |
213 |
12 |
9 |
662 |
154 |
176 |
10 |
9 |
169 |
46 |
46 |
5 |
SecJov |
4 |
649 |
82 |
153 |
17 |
4 |
332 |
43 |
88 |
14 |
- |
- |
- |
- |
- |
PasInv |
11 |
1640 |
104 |
104 |
8 |
11 |
359 |
57 |
57 |
6 |
- |
- |
- |
- |
- |
PasLim |
9 |
1534 |
74 |
84 |
7 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
Travessão338S |
|||||||||||||||
FloCon |
7 |
990 |
173 |
277 |
19 |
7 |
530 |
105 |
162 |
14 |
7 |
117 |
41 |
53 |
6 |
FloExp |
16 |
1885 |
333 |
333 |
14 |
16 |
996 |
177 |
177 |
10 |
16 |
226 |
59 |
59 |
4 |
SecAnt |
4 |
307 |
81 |
218 |
31 |
4 |
392 |
93 |
265 |
34 |
4 |
64 |
31 |
77 |
18 |
SecJov |
5 |
624 |
96 |
180 |
21 |
5 |
958 |
68 |
134 |
18 |
5 |
6 |
7 |
13 |
6 |
PlaArb |
4 |
706 |
91 |
187 |
25 |
4 |
245 |
42 |
106 |
21 |
4 |
23 |
11 |
31 |
12 |
PasInv |
6 |
990 |
56 |
107 |
15 |
6 |
42 |
13 |
30 |
8 |
6 |
7 |
3 |
4 |
2 |
-----

Figura 2. Curvas de acumulação de riqueza de espécies por abundância de indivíduos que comparam os tipos de uso da terra (ver siglas na Tabela 1) por estrato (inferior, médio, superior) em três mosaicos rurais (Palmares II, Maçaranduba, Travessão 338-S) na região do “arco do desmatamento”, Amazônia Oriental. Linhas contínuas indicam interpolação e linhas tracejadas indicam extrapolação. Letras diferentes indicam diferença significativa entre tipos de uso considerando os intervalos de confiança a 95% (áreas sombreadas).
A ausência de fragmentos florestais conservados e o efeito do fogo nas florestas queimadas de Palmares II pode ter contribuído para uma menor riqueza das florestas secundárias antigas nos estratos inferior e médio deste mosaico. Sabemos que fragmentos florestais conservados inseridos nos mosaicos servem como importantes fontes de dispersão (Vieira e Proctor 2007); por outro lado, o fogo altera drasticamente a composição florística, reduz o dossel florestal, biomassa e número de árvores adultas (Barlow e Peres, 2008). O efeito do fogo no dossel florestal reduziu o número de indivíduos nas florestas queimadas de Palmares II em comparação com as florestas exploradas, tanto no estrato superior quanto no médio (Figura 2b-c; Tabela 2)
Como em Palmares II, as florestas secundárias não tiveram riqueza similar às florestas nos estratos inferior e médio, e as florestas queimadas foram drasticamente alteradas pelo fogo nos estratos médio e superior, apenas as florestas exploradas podem ser consideradas como coberturas florestais funcionais (CFF), ou seja, aqueles fragmentos de alta diversidade, capazes de promover o funcionamento ecossistêmico da paisagem. Esse resultado associado ao menor número de pontos amostrais em florestas exploradas (cinco pontos), revela quão frágil é o ambiente em Palmares II, pois alguns estudos mostram que quanto maior a biodiversidade dos fragmentos florestais, maior é a capacidade do ecossistema de exercer funções como produção de biomassa, decomposição e absorção de nutrientes, que irão influenciar diretamente na manutenção de serviços ecossistêmicos como estoque de carbono, produção de serrapilheira e produção de madeira (Ferraz et al., 2014; Isbell et al., 2014).
Evidências teóricas e empíricas sugerem que a persistência da biodiversidade nas paisagens alteradas declina acentuadamente quando a cobertura florestal corresponde a menos de 30% da paisagem (Banks-Leite et al., 2014). As florestas exploradas, únicas CFF em Palmares II, correpondem a 11% dos pontos amostrais. Em Maçaranduba e Travessão 338-S, a porcentagem de CFF é maior (44 e 60%, respectivamente), pois além das florestas conservadas e exploradas, as florestas secundárias antigas também possuem elevada riqueza de espécies. Portanto, a menor porcentagem de CFF em Palmares II tem forte contribuição sobre a menor riqueza encontrada nas florestas secundárias antigas deste mosaico.
A falta de CFF em Palmares II reflitiu não apenas na regeneração das florestas secundárias antigas, mas também nas secundárias jovens, as quais tiveram riqueza similar aos cultivos anuais e pastos invadidos nos estratos inferior e médio (Figura 2a-b; Tabela 2). As florestas secundárias jovens e os plantios arbóreos (esses últimos presentes apenas no Travessão 338-S), tiveram riqueza intermediárias entre os cultivos agrícolas e as florestas conservadas, exploradas e secundárias antigas em Maçaranduba e Travessão 338-S (Figura 2d-i; Tabela 2). Os plantios arbóreos no Travessão 338-S tiveram até riqueza similar às florestas conservadas, exploradas e secundárias antigas no estrato superior, mas com baixo número de indivíduos (Figura 2i; Tabela 2). A maior riqueza nos plantios arbóreos (ao contrário dos pastos invadidos) se deve ao fato de promoverem condições ambientais propícias à regeneração de árvores, através do estabelecimento de um microclima mais favorável e a atração de agentes dispersores (Carnevale e Montagnini, 2002; Cusack e Montagnini, 2004). O mesmo acontece nas florestas secundárias, que desempenham um grande papel na conservação das espécies na paisagem.
O aumento de florestas secundárias na paisagem pode contribuir para manter mais espécies através do incremento da cobertura florestal na escala da paisagem (Gardner et al., 2008; Arroyo-Rodriguez et al., 2015). Além disso, as múltiplas vias da sucessão que as florestas secundárias podem seguir implicam em maior diversidade beta, permitindo assim a acumulação de um maior número de espécies nos mosaicos rurais (Arroyo-Rodriguez et al., 2013; Puttker et al., 2015).
Finalmente, a menor riqueza encontrada nos pastos é atribuída à regeneração limitada nesses ambientes, principalmente por dois motivos (Elgar et al. 2014): (1) falta de viabilidade dos propágulos florestais, seja pela exaustão do banco de sementes durante sucessivos cultivos ou pela ausência de animais dispersores nos pastos; e, (2) pela competição que desempenha um papel importante, uma vez que as espécies forrageiras nas pastagens, como a Urochloa spp., cultivadas nas pastagens dos mosaicos estudados, alteram as condições microclimáticas, umidade e nutrientes do solo (Cheung et al., 2009; Shoo e Catterall, 2013).
O inventário florístico mostrou que os números de espécies diminuem do chão (estrato inferior) para o dossel (estrato superior). O estrato inferior de todos os mosaicos apresentou, em média, 5,5 vezes mais espécies que o estrato superior e, o estrato médio, por sua vez, apresentou 3 vezes mais espécies que o estrato superior (Tabela 3).
Os estratos inferiores podem corresponder de 21 a 47% da riqueza total de espécies em florestas tropicais (Gentry e Dodson, 1987). Em florestas secundárias, essa riqueza tende a aumentar à medida que o tempo de paralisação das atividades agrícolas avança (Chazdon, 2012). Outro fator que contribui para a maior riqueza nos estratos inferiores é a alta taxa de mudanças na composição florística (Costa et al. 2005), causada pela rápida entrada de espécies pioneiras nas comunidades perturbadas, pela permanência de algumas populações florestais que ainda se regeneram nos ambientes agrícolas e, por meio do banco de sementes e rebrotos (Vieira e Proctor, 2007; Mitja et al., 2008; Miranda et al., 2009).
Tabela 3. Número de indivíduos (Ind), espécies inventariadas (S), número estimado de espécies (S-est) e desvio padrão estimado (SD) encontrados nos três estratos florísticos (inferior, médio, superior) de três mosaicos rurais (Palmares II, Maçaranduba, Travessão 338-S) na região do “arco do desmatamento”, Amazônia Oriental. Número estimado de espécies e desvio padrão estimado foram obtidos a partir da extrapolação do número de amostras considerando n = 45.
Mosaicos |
n |
Ind |
S |
S est |
SD |
Estrato Inferior |
|||||
Palmares II |
45 |
10071 |
511 |
511 |
17 |
Maçaranduba |
45 |
6643 |
438 |
438 |
13 |
Travessão 338S |
45 |
6229 |
542 |
542 |
15 |
Estrato Médio |
|||||
Palmares II |
38 |
6270 |
253 |
276 |
12 |
Maçaranduba |
32 |
2316 |
316 |
369 |
13 |
Travessão 338S |
41 |
3290 |
315 |
331 |
13 |
Estrato Superior |
|||||
Palmares II |
23 |
224 |
88 |
124 |
11 |
Maçaranduba |
20 |
430 |
95 |
140 |
13 |
Travessão 338S |
37 |
444 |
88 |
95 |
5 |
Houveram diferenças significativas entre as curvas de rarefação de espécies entre os mosaicos nos estratos inferior e médio (Figura 3; Tabela 3). No estrato inferior, Travessão 338-S (542 ± 15 esp.; número de espécies e desvio padrão estimados) apresentou mais espécies que Palmares II (511 ± 17 esp.) e Maçaranduba (438 ± 13 esp.). No estrato médio, Maçaranduba (369 ± 13 esp.) teve maior riqueza que Travessão 338-S (331 ± 13 esp.), e ambos os mosaicos tiveram maior riqueza que Palmares II (276 ± 12 esp.). No estrato superior, não houve diferença significativa entre a riqueza de espécies dos mosaicos, apesar de Maçaranduba ter mostrado mais indivíduos (Figura 3; Tabela 3).
A menor riqueza no estrato inferior de Maçaranduba e Palmares II pode ser consequência da maior concentração de pastos e cultivos nestes mosaicos. Como dito anteriormente, as pastagens e os cultivos apresentaram a menor riqueza de espécies entre os tipos de uso estudados. Competição com forrageiras e fonte de dispersão limitada, associadas com o preparo de área e o manejo do gado em pastagens, são as principais causas da baixa regeneração nos tipos de uso agrícolas (Esquivel et al. 2008; Mitja et al. 2008; Elgar et al. 2014).

Figura 3. Curvas de acumulação de espécies por amostra que comparam os três estratos florísticos (a = inferior, b = médio e c = superior) entre três mosaicos rurais (Palmares II, Maçaranduba, Travessão 338-S) na região do “arco do desmatamento”, Amazônia Oriental. Linhas contínuas indicam interpolação e linhas tracejadas indicam extrapolação. Letras diferentes indicam diferença significativa entre mosaicos considerando os intervalos de confiança a 95% (áreas sombreadas por tipo de uso).
Por outro lado, Maçaranduba apresentou a maior riqueza entre os mosaicos no estrato médio, o que indica maior estabelecimento de espécies lenhosas na paisagem, possivelmente em resposta ao maior número de florestas secundárias neste mosaico (13 pontos amostrais) em comparação com Travessão 338-S e Palmares II (nove pontos cada). A regeneração florestal pode avançar rapidamente e acumular espécies a taxas elevadas em algumas florestas tropicais (Lebrija-Trejos et al. 2010), contudo, essa capacidade de resiliência ecológica das florestas secundárias é maior e mais previsível em paisagens onde florestas nativas bem preservadas, ricas em biodiversidade ainda estão presentes na paisagem (Jakovac et al., 2015; Reyer et al. 2015). Maçaraduba e Travessão 338-S ainda possuem florestas conservadas, e por isso tiveram mais espécies no estrato médio que Palmares II; contudo o Travessão 338-S possui maior cobertura florestal que Maçaranduba. Paisagens com cobertura florestal intermediária (ou seja, 20-50%), como em Maçaranduba, mostram maior variabilidade no grau de fragmentação e na borda total da floresta (Villard e Metzger, 2014), portanto, as diferenças na estrutura da paisagem e na dinâmica do uso da terra podem conduzir a diversas vias de regeneração (Arroyo-Rodriguez, et al. 2015; Jakovac et al., 2015).
Esse caráter intermediário de Maçaranduba, nos remete à hipótese dos distúrbios intermediários, normalmente aplicados às florestas primárias (Sheil e Burslem 2003); essa hipótese explica a maior riqueza quando espécies de diferentes estádios sucessionais convivem simultaneamente. Espécies pioneiras foram associadas às florestas de Maçaranduba, enquanto esse padrão não foi encontrado no Travessão 338-S (Do Vale, dados não publicados).
Apesar das diferenças nos estratos inferior e médio, não houve diferença na riqueza do estrato superior entre os mosaicos; entretanto, após considerar as diferentes paisagens estabelecidas em cada mosaico, principalmente relacionadas a quantidade de CFF presente, é provável que haja alta heterogeneidade na composição e dominância florística do estrato superior entre os mosaicos. Na Mata Atlântica, em paisagens de intensa fragmentação, ocorre uma redução na riqueza de espécies (em até 50%) e maior de dominância de espécies pioneiras, ou “secundarização” dos fragmentos florestais (Joly et al. 2014). Essas assembléias dominadas por pioneiras podem persistir como matrizes e permanecer dominados por habitats abertos, como pastagens (Tabarelli et al., 2008).
A diversidade beta entre fazendas contribuiu mais fortemente para a biodiversidade total nos mosaicos rurais, relacionada com a composição de tipos de uso implantados em cada fazenda, e tende a aumentar com a fragmentação dos tipos de uso da terra na paisagem.
A riqueza das florestas secundárias depende da quantidade de fragmentos florestais de alta riqueza dentro do mosaico; se existem muitos fragmentos florestais, então a riqueza das florestas secundárias antigas será alta, similar à riqueza das florestas; se existem poucos fragmentos florestais, ou alta degradação, como florestas queimadas, então a riqueza das florestas secundárias antigas será similar aos cultivos agrícolas, ou às florestas degradadas.
Os padrões de riqueza foram diferentes entre os estratos, à medida que a inclusão dos estratos inferior e médio ajudaram a acessar diferenças na riqueza de espécies entre tipos de uso florestais e agrícolas. A menor riqueza no estrato inferior dos mosaicos foi associada a maior quantidade de tipos de uso agrícolas, enquanto a maior riqueza no estrato médio foi associada a maior quantidade de florestas secundárias.
Este estudo foi financiado pela Agence Nationale de la Recherche, da França, através dos processos ANR-06-PADD-001-011 and ANR 06 BIODIV 009-01 (IFB-ANR); pelo governo brasileiro através do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e Instituto Nacional de Ciência e Tecnologia – Biodiversidade e Uso da Terra na Amazônia. O CNPq também concedeu a bolsa de estudo a Igor Do Vale. Nós agradecemos aos pequenos produtores de Maçaranduba, Palmares II and Travessão 338-S pela disponibilidade e pela ajuda no campo, ao Sr. Deurival da Costa Carvalho pela sua eficiência e entusiasmo durante o trabalho de campo; e aos alunos da Universidade Federal Rural da Amazônia Livia Teixeira, Luciana Brabo, Helber Shibata, Fábio Leão, Joice Silva, José Moreira, Heraldo Silva, Robson Ramos e Magno Reis por nos ajudarem no trabalho de campo.
Arroyo‐Rodríguez, V., Rös, M., Escobar, F., Melo, F. P., Santos, B. A., Tabarelli, M., & Chazdon, R. (2013). Plant β‐diversity in fragmented rain forests: testing floristic homogenization and differentiation hypotheses. Journal of Ecology, 101(6), 1449-1458.
Arroyo‐Rodríguez, V., Melo, F. P., Martínez‐Ramos, M., Bongers, F., Chazdon, R. L., Meave, J. A., ... & Tabarelli, M. (2015). Multiple successional pathways in human‐modified tropical landscapes: new insights from forest succession, forest fragmentation and landscape ecology research. Biological Reviews. doi:10.1111/brv.12231
Banks-Leite, C., Pardini, R., Tambosi, L. R., Pearse, W. D., Bueno, A. A., Bruscagin, R. T., ... & Metzger, J. P. (2014). Using ecological thresholds to evaluate the costs and benefits of set-asides in a biodiversity hotspot. Science, 345(6200), 1041-1045.
Barlow, J., & Peres, C. A. (2008). Fire-mediated dieback and compositional cascade in an Amazonian forest. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 363(1498), 1787-1794.
Benton, T. G., Vickery, J. A., & Wilson, J. D. (2003). Farmland biodiversity: is habitat heterogeneity the key?. Trends in Ecology & Evolution, 18(4), 182-188.
Carnevale, N. J., & Montagnini, F. (2002). Facilitating regeneration of secondary forests with the use of mixed and pure plantations of indigenous tree species. Forest ecology and management, 163(1), 217-227.
Carvalho, J., Marques, M. C. M., Roderjan, C. V., Barddal, M., & Sousa, S. D. (2009). Relações entre a distribuição das espécies de diferentes estratos e as características do solo de uma floresta aluvial no Estado do Paraná, Brasil. Acta Botanica Brasilica, 23(1), 1-9.
Chazdon, R. (2012). Regeneração de florestas tropicais Tropical forest regeneration. Boletim Museu Paraense Emílio Goeldi de Ciencias Naturais, 7, 195-218.
Cheung, K. C., Marques, M. C., & Liebsch, D. (2009). Relação entre a presença de vegetação herbácea e a regeneração natural de espécies lenhosas em pastagens abandonadas na Floresta Ombrófila Densa do Sul do Brasil. Acta botanica brasilica, 23(4), 1048-1056.
Chisholm, R. A., Muller‐Landau, H. C., Abdul Rahman, K., Bebber, D. P., Bin, Y., Bohlman, S. A., ... & Cao, H. (2013). Scale‐dependent relationships between tree species richness and ecosystem function in forests. Journal of Ecology, 101(5), 1214-1224.
Colwell, R. K., Chao, A., Gotelli, N. J., Lin, S. Y., Mao, C. X., Chazdon, R. L., & Longino, J. T. (2012). Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. Journal of plant ecology, 5(1), 3-21.
Colwell, R. K. 2013. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9 and earlier. User’s guide and application. Published at: http://purl.oclc.org/estimates
Costa, F. R., Magnusson, W. E., & Luizao, R. C. (2005). Mesoscale distribution patterns of Amazonian understorey herbs in relation to topography, soil and watersheds. Journal of Ecology, 93(5), 863-878.
Costa, L. G. S., Miranda, I. S., Grimaldi, M., Silva, M. L., Mitja, D., & Lima, T. T. S. (2012). Biomass in different types of land use in the Brazil’s ‘arc of deforestation’. Forest Ecology and Management, 278, 101-109.
Crist, T. O., Veech, J. A., Gering, J. C., & Summerville, K. S. (2003). Partitioning species diversity across landscapes and regions: a hierarchical analysis of α, β, and γ diversity. The American Naturalist, 162(6), 734-743.
Cusack, D., & Montagnini, F. (2004). The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. Forest Ecology and Management, 188(1), 1-15.
De Castro Solar, R. R., Barlow, J., Andersen, A. N., Schoereder, J. H., Berenguer, E., Ferreira, J. N., & Gardner, T. A. (2016). Biodiversity consequences of land-use change and forest disturbance in the Amazon: a multi-scale assessment using ant communities. Biological Conservation, 197, 98-107.
De Sartre, X. A., Oszwald, J., Ramírez, B., Rodríguez, G., Velazquez, J., Sebille, P., & Lavelle, P. (2011). Caractéristiques socio-économiques d'une intégration régionale en Amazonie colombienne (région de Florencia, Caqueta). Caravelle (1988-), 91-110.
Do Vale, I., Miranda, I. S., Mitja, D., Grimaldi, M., Nelson, B. W., Desjardins, T., & Costa, L. G. S. (2015). Tree Regeneration Under Different Land-Use Mosaics in the Brazilian Amazon’s “Arc of Deforestation”. Environmental management, 56(2), 342-354.
Elgar, A. T., Freebody, K., Pohlman, C. L., Shoo, L. P., & Catterall, C. P. (2014). Overcoming barriers to seedling regeneration during forest restoration on tropical pasture land and the potential value of woody weeds. Frontiers in plant science, 5. doi: 10.3389/fpls.2014.00200
Esquivel, M. J., Harvey, C. A., Finegan, B., Casanoves, F., & Skarpe, C. (2008). Effects of pasture management on the natural regeneration of neotropical trees. Journal of applied ecology, 45(1), 371-380.
Ferraz, S. F., Ferraz, K. M., Cassiano, C. C., Brancalion, P. H. S., da Luz, D. T., Azevedo, T. N.,... & Metzger, J. P. (2014). How good are tropical forest patches for ecosystem services provisioning?. Landscape ecology, 29(2), 187-200.
Fortin, M. J., Dale, M. R. (2005). Spatial analysis: a guide for ecologists. Cambridge University Press, Cambridge, UK.
Gardner, T. A., Barlow, J., Parry, L. W., & Peres, C. A. (2007). Predicting the uncertain future of tropical forest species in a data vacuum. Biotropica, 39(1), 25-30.
Gardner, T. A., Hernández, M. I., Barlow, J., & Peres, C. A. (2008). Understanding the biodiversity consequences of habitat change: the value of secondary and plantation forests for neotropical dung beetles. Journal of applied ecology, 45(3), 883-893.
Gentry, A. H., & Dodson, C. (1987). Contribution of nontrees to species richness of a tropical rain forest. Biotropica, 149-156.
Gibson, L., Lee, T. M., Koh, L. P., Brook, B. W., Gardner, T. A., Barlow, J., ... & Sodhi, N. S. (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature, 478(7369), 378-381.
Grimaldi, M., Oszwald, J., Dolédec, S., del Pilar Hurtado, M., de Souza Miranda, I., De Sartre, X. A., ... & Guevara, E. (2014). Ecosystem services of regulation and support in Amazonian pioneer fronts: searching for landscape drivers. Landscape ecology, 29(2), 311-328.
Isbell, F., Tilman, D., Polasky, S., & Loreau, M. (2014). The biodiversity‐dependent ecosystem service debt. Ecology letters, 18(2), 119-134.
Jakovac, C. C., Peña‐Claros, M., Kuyper, T. W., & Bongers, F. (2015). Loss of secondary‐forest resilience by land‐use intensification in the Amazon. Journal of Ecology, 103(1), 67-77.
Kennard, D. K. (2002). Secondary forest succession in a tropical dry forest: patterns of development across a 50-year chronosequence in lowland Bolivia. Journal of tropical ecology, 53-66.
Kessler, M., Abrahamczyk, S., Bos, M., Buchori, D., Putra, D. D., Gradstein, S. R., ... & Saleh, S. (2009). Alpha and beta diversity of plants and animals along a tropical land‐use gradient. Ecological Applications, 19(8), 2142-2156.
Lande, R. (1996). Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos, 5-13.
Lavelle, P., Dolédec, S., De Sartre, X. A., Decaëns, T., Gond, V., Grimaldi, M., ... & De Souza, S. (2016). Unsustainable landscapes of deforested Amazonia: An analysis of the relationships among landscapes and the social, economic and environmental profiles of farms at different ages following deforestation. Global Environmental Change, 40, 137-155.
Lebrija-Trejos, E., Meave, J. A., Poorter, L., Pérez-García, E. A., & Bongers, F. (2010). Pathways, mechanisms and predictability of vegetation change during tropical dry forest succession. Perspectives in Plant Ecology, Evolution and Systematics, 12(4), 267-275.
Marichal, R., Grimaldi, M., Feijoo, A., Oszwald, J., Praxedes, C., Cobo, D. H. R., ... & Miranda, I. S. (2014). Soil macroinvertebrate communities and ecosystem services in deforested landscapes of Amazonia. Applied Soil Ecology, 83, 177-185.
Melo, F. P., Arroyo-Rodríguez, V., Fahrig, L., Martínez-Ramos, M., & Tabarelli, M. (2013). On the hope for biodiversity-friendly tropical landscapes. Trends in Ecology & Evolution, 28(8), 462-468.
Mendes, R. S., Evangelista, L. R., Thomaz, S. M., Agostinho, A. A., & Gomes, L. C. (2008). A unified index to measure ecological diversity and species rarity. Ecography, 31(4), 450-456.
Mesquita, R. D. C. G., dos Santos Massoca, P. E., Jakovac, C. C., Bentos, T. V., & Williamson, G. B. (2015). Amazon rain forest succession: stochasticity or land-use legacy?. Bioscience, 65(9), 849-861.
Miranda, I. S., Mitja, D., Silva, T. S. (2009). Mutual influence of forests and pastures on the seedbanks in the Eastern Amazon. Weed Research, 49, 499-505.
Mitja, D., de Souza Miranda, I., Velasquez, E., & Lavelle, P. (2008). Plant species richness and floristic composition change along a rice-pasture sequence in subsistence farms of Brazilian Amazon, influence on the fallows biodiversity (Benfica, State of Pará). Agriculture, ecosystems & environment, 124(1), 72-84.
Morandi, P. S., Marimon, B. S., Eisenlohr, P. V., Marimon-Junior, B. H., Oliveira-Santos, C., Feldpausch, T. R., ... & Phillips, O. L. (2016). Patterns of tree species composition at watershed-scale in the Amazon ‘arc of deforestation’: implications for conservation. Environmental Conservation, 1-10.
Newbold, T., Hudson, L. N., Hill, S. L., Contu, S., Lysenko, I., Senior, R. A., ... & Day, J. (2015). Global effects of land use on local terrestrial biodiversity. Nature, 520(7545), 45-50.
Oszwald, J., Gond, V., Doledec, S., & Lavelle, P. (2011). Identification d’indicateurs de changement d’occupation du sol pour le suivi des mosaïques paysagères. Bois et forêts des tropiques, 307(1), 7-21.
Oszwald, J., De Sartre, X. A., Decaens, T., Gond, V., Grimaldi, M., Lefebvre, A., … Lavelle, P. (2012). Utilisation de la télédétection et de données socio-économiques et écologiques pour comprendre l'impact des dynamiques de l'occupation des sols à Pacajà (Brésil). Revue Française de Photogrammétrie et de Télédétection, 198-199, 8-24.
Pausas, J. G., & Austin, M. P. (2001). Patterns of plant species richness in relation to different environments: an appraisal. Journal of Vegetation Science, 12(2), 153-166.
Puig, H. 2008. A floresta tropical úmida. Editora UNESP, São Paulo. 496p.
Püttker, T., de Arruda Bueno, A., Prado, P. I., & Pardini, R. (2015). Ecological filtering or random extinction? Beta‐diversity patterns and the importance of niche‐based and neutral processes following habitat loss. Oikos, 124(2), 206-215.
Reyer, C. P., Brouwers, N., Rammig, A., Brook, B. W., Epila, J., Grant, R. F., ... & Medlyn, B. (2015). Forest resilience and tipping points at different spatio‐temporal scales: approaches and challenges. Journal of Ecology, 103(1), 5-15.
Sfair, J. C., Arroyo‐Rodríguez, V., Santos, B. A., & Tabarelli, M. (2016). Taxonomic and functional divergence of tree assemblages in a fragmented tropical forest. Ecological Applications, 26(6), 1816-1826.
Sheil, D., & Burslem, D. F. (2003). Disturbing hypotheses in tropical forests. Trends in Ecology & Evolution, 18(1), 18-26.
Shoo, L. P., & Catterall, C. P. (2013). Stimulating natural regeneration of tropical forest on degraded land: approaches, outcomes, and information gaps. Restoration Ecology, 21(6), 670-677.
Tabarelli, M., Lopes, A. V., & Peres, C. A. (2008). Edge‐effects Drive Tropical Forest Fragments Towards an Early‐Successional System. Biotropica, 40(6), 657-661.
Tscharntke, T., Tylianakis, J. M., Rand, T. A., Didham, R. K., Fahrig, L., Batary, P., ... & Ewers, R. M. (2012). Landscape moderation of biodiversity patterns and processes‐eight hypotheses. Biological Reviews, 87(3), 661-685.
Tylianakis, J. M., Klein, A. M., & Tscharntke, T. (2005). Spatiotemporal variation in the diversity of Hymenoptera across a tropical habitat gradient. Ecology, 86(12), 3296-3302.
Van Breugel, M., Hall, J. S., Craven, D., Bailon, M., Hernandez, A., Abbene, M., & van Breugel, P. (2013). Succession of ephemeral secondary forests and their limited role for the conservation of floristic diversity in a human-modified tropical landscape. PLoS One, 8(12), e82433.
Veech, J. A., Summerville, K. S., Crist, T. O., & Gering, J. C. (2002). The additive partitioning of species diversity: recent revival of an old idea. Oikos, 99(1), 3-9.
Vieira, I. C. G., & Proctor, J. (2007). Mechanisms of plant regeneration during succession after shifting cultivation in eastern Amazonia. Plant Ecology, 192(2), 303-315.
Villard, M. A., & Metzger, J. P. (2014). REVIEW: beyond the fragmentation debate: a conceptual model to predict when habitat configuration really matters. Journal of Applied Ecology, 51(2), 309-318.
1. Engenheiro Florestal, Doutorando no Programa de Pós-Graduação em Ciências Florestais, Universidade Federal Rural da Amazônia. dovale.igor@gmail.com
2. Bióloga, Professora do Instituto Socioambiental e dos Recursos Hídricos, Universidade Federal Rural da Amazônia. izildinha.miranda@ufra.edu.br
3. Bióloga, Pesquisadora do Institut de Recherche pour le Développement. danielle.mitja@ird.fr
4. Biólogo, Doutorando no Programa de Pós-Graduação em Ciências Florestais, Universidade Federal Rural da Amazônia. ggaldino@gmail.com
5. Agrônomo, Universidade Federal do Sul e Sudeste do Pará. Agrônomo. alessio@ufpa.br
6. Engenheira Florestal, Professora do Campus Paragominas, Universidade Federal Rural da Amazônia. tamara.lima@ufra.edu.br
7. Engenheiro Florestal, Professor do Instituto Socioambiental e dos Recursos Hídricos, Universidade Federal Rural da Amazônia. luizgonzagacosta53@gmail.com