![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 31) Año 2017. Pág. 3
Alberto DORVAL 1; Otávio PERES Filho 2; Valdiclei Custódio JORGE 3; Marcelo Dias de SOUZA 4; Wilian Oliveira ROCHA 5
Recibido: 17/01/2017 • Aprobado: 03/03/2017
RESUMO: Este trabalho teve como objetivo conhecer a diversidade de formigas em área de cerradão com aproximadamente 20 hectares com e sem exploração de madeira. O trabalho foi realizado de setembro de 2011 a agosto de 2012, no município de Cuiabá, MT, utilizando 32 armadilhas, tipo pitfall, constituída de pote plástico de 400ml contendo água + sal + detergente neutro, cujo as coletas foram quinzenais. Foram amostradas duas áreas: (A) - Área com extração de madeira (1,5 ha) e (B) - Área com vegetação preservada (1,5 ha), utilizando 16 armadilhas pitfall distribuídas em duas linhas para cada área, sendo oito armadilhas por linha e uma distância de 2 metros entre armadilhas e 10 metros entre linhas. Coletou-se 16.373 indivíduos, sendo 10.260 na área com exploração de madeira e 6.113 na área sem exploração de madeira, distribuídos em cinco subfamílias, 17 gêneros e 25 espécies. Myrmicinae, Formicinae e Ecitoninae foram as subfamílias mais diversificadas e abundantes. A riqueza de espécies não foi afetada pela extração de madeira, e sim, a composição e densidade populacional das espécies, principalmente em relação as espécies mais adaptadas aos ambientes antropizados. As espécies Camponotus rufipes, Pheidole rodeszkowskii, Solenopsis invicta na área explorada, Labidus praedator e Pheidole fimbriata na área preservada foram as mais representativas em número de indivíduos. |
ABSTRACT: This work had as objective knows the diversity of ants in savanna area with approximately 20 hectares with and without wood exploration. The work was accomplished of September of 2011 to August of 2012, in the municipal district of Cuiabá, MT, using 32 traps, type pitfall, constituted of plastic pot of 400 ml containing water + salt + neutral detergent, being the collects biweekly. Were sampled two areas: (A) - Area with wood extraction (1,5 ha) and (B) - Area with preserved vegetation (1,5 ha). In each area were utilizing 16 pitfall traps distributed in two lines, with eight traps per line and distance of 2 meters between traps and 10 meters between lines. Was collected 16.373 individuals, being 10.260 in the area with wood exploration and 6.113 in the area without wood exploration, distributed in 5 subfamilies, 17genera, 25 species. Myrmicinae, Formicinae and Ecitoninae were the subfamilies more diversified and abundant. The diversity of species was not affected by the wood extraction, however the composition and the population density of the species was different, Especially in relation to the species most adapted to the anthropized environments. The species Camponotus rufipes, Pheidole rodeszkowskii, Solenopsis invicta in the explored area, Labidus praedator and Pheidole fimbriata in the preserved area, were the most representative in number of individuals. |

A cidade de Cuiabá é conhecida como “cidade-verde” devido à abundância de áreas com vegetação de cerrado e cerradão localizados dentro da área urbana. Porém, nas ultimas décadas, estes remanescentes florestais nativos tem sofrido uma redução devido ao avanço do processo de urbanização, que tem contribuído para sua fragmentação.
Em fragmentos urbanos, fatores antrópicos como o constante avanço da urbanização, grilagem de terra por posseiros e grileiros profissionais, mineração, extração ilegal de madeira, de cascalho, plantios de culturas agrícolas e pastagens são os responsáveis pelo empobrecimento da biodiversidade e a extinção da flora e fauna dentro destes remanescentes florestais nativos. Portanto, ações urgentes devem ser direcionadas para conhecer o impacto destas atividades sobre diversidade e abundância da entomofauna nativa que ainda ocorrem nestes fragmentos.
Uma das maneiras de detectar e monitorar os padrões de mudança na biodiversidade provocados por ações humanas é utilizar espécies, ou mesmo grupo de espécies, que funcionam como bioindicadoras da degradação ambiental (SANTOS et al., 2006). Um exemplo de espécies bioindicadora são as formigas, pois os efeitos destas perturbações antrópicas sobre as comunidades arborícolas influenciam diretamente na mirmecofauna, causando perda da diversidade, mudanças na composição de espécies, a alteração das interações interespecíficas, mudanças nas relações trófica, nas interações formigas-plantas, entre formigas e hemípteros sugadores de seiva, modificação nas taxas de dispersão de sementes e de predação, além da modificação do solo (PHILPOTT et al., 2010).
As formigas por constituírem um grupo de insetos de hábito social com uma ampla distribuição na região tropical (WILSON, 1971), colonizando diferentes ambientes, além de apresentarem fidelidade ecológica ao seu local de ocorrência natural, têm sido utilizadas para medir o efeito de diferentes atividades antrópicas na diversidade e abundância dentro de fragmentos florestais nativos.
Este grupo de insetos caracteriza-se pela alta diversidade e susceptibilidade às mudanças antrópicas, apresentando potencial para serem utilizadas em estudos de impacto ambiental, pois são abundantes, diversificados em espécies, ampla distribuição geográfica, facilidade de amostragem e de identificação taxonômica (SILVA; BRANDÃO, 1999, ALONSO; DONATI, 2000). De acordo com Santos et al. (2006), mudanças nas características ambientais dos fragmentos podem influenciar na riqueza local de espécies de formigas, pois estas dependem da diversidade de habitats disponíveis, como a estrutura física e a quantidade de serapilheira, onde encontram recursos alimentares e sítios de nidificação utilizados por diversas espécies de formigas.
A escolha do método de amostragem utilizado para monitorar formigas deve ser eficiente e apresentar um baixo custo/benefício e a armadilha do tipo pitfall é um dos métodos mais simples para amostrar artrópodes de serapilheira, pois apresenta vantagens, como uso por longos períodos e serem eficientes na coleta de espécies predadoras e saprófagas (OLSON, 1991). De acordo com Freitas et al. (2004) e Vargas et al.(2007) as armadilhas do tipo pitfall são as mais indicadas e ideais para coletas de formigas, principalmente em fitofisionomia de cerrado.
Estudos com formigas, com a finalidade de medir impacto e recuperações ambientais causada por atividades antrópicas foram conduzidas por Majer (1983, 1992, 1996), Majer e Nichols (1998), Lobry de Bruyn (1999), Ketelhut (1999), Brandão et al. (2000), Vasconcelos et al. (2000), Corrêa et al.(2006), Campos et al. (2008), Soares et al.(2010), Gunawardene et. al. (2010), Freire et al.( 2012), Suguituru et al.(2013), Dittmann et al. (2014), Rocha et al. (2015), Sousa et al. (2015). Portanto o presente estudo teve como objetivo conhecer a riqueza e abundância de Formicidae em uma área com cobertura florestal inalterada e outra antropizada pela exploração de madeira em um fragmento de cerradão, no perímetro urbano, no município de Cuiabá, MT.
O trabalho foi realizado de setembro de 2011 a agosto de 2012, no Bairro Coxipó-da-Ponte, no município de Cuiabá, estado de Mato Grosso, em um fragmento florestal, tipo cerradão, também denominado de Cerrado fechado, pois possui aspecto florestal devido a grande quantidade de árvores e formação de dossel (SANTOS, 2014). A área de estudo de aproximadamente 20 hectares, localiza-se sob as coordenadas 15°36'37"S e 56°3'47"W.
O solo é podzólico vermelho-amarelo distrófico, com média a baixa fertilidade. A altitude média é de 165 metros em relação ao nível do mar. O clima é tropical quente e sub-úmido com temperatura média anual de 27°C e precipitação pluvial anual de 1350 mm. Dentro do fragmento florestal foram amostradas duas áreas: (A) - Área com extração de madeira (1,5 ha), sendo esta área caracterizada pela extração de madeira de forma tradicional e rudimentar, sem a utilização de qualquer tipo de manejo florestal, para o uso como lenha, na fabricação de carvão e mourões de cerca; (B)- Área com vegetação preservada (1,5 ha) sem nenhuma intervenção humana.
As armadilhas utilizadas foram do tipo pitfall composta de um pote de plástico de 15 cm de altura e 2,5cm de diâmetro, enterradas com suas bordas ao nível do solo e preenchidas com solução conservante contendo água + detergente neutro + sal, a um terço da capacidade do recipiente (Figura 1). Dentro de cada área foram utilizadas 16 armadilhas pitfall distribuídas em duas linhas paralela entre si, contendo oito armadilhas cada, sendo os distanciamento entre as armadilhas e as linhas, com 2 m e 10 m respectivamente.

Figura 1. Armadilha do tipo pitfall utilizado no levantamento
da mirmecofauna. Cuiabá, Mato Grosso, Brasil, 2011.
As coletas foram realizadas quinzenalmente e os espécimes coletados, foram individualizados por área e armadilha, da qual foram armazenados e etiquetados com as características do local, sendo encaminhados para o Laboratório de Proteção Florestal da Faculdade de Engenharia Florestal, da Universidade Federal de Mato Grosso para triagem e quantificação. Posteriormente os exemplares foram enviados para o Laboratório de Mirmecologia do CEPLAC-CEPEC, Ilhéus, Bahia, sendo identificados pelo Prof. Dr. Jacques H. C. Delabie.
Os resultados foram tabulados e analisados quantitativamente e qualitativamente de acordo com ocorrência dos formicídeos em relação ao meio ambiente analisado, levando em consideração os períodos climáticos. O índice de diversidade foi calculado através do Programa Statistical Ecology, utilizando os números de HILL (N0, N1 e N2) por serem de fácil interpretação e tem a vantagem de sua unidade de medida ser expressa em espécie (Ludwig & Reynolds, 1988).
Para relatar alteração de ocorrência dos formicídeos em função dos períodos climáticos, os dados foram analisados utilizando técnicas de estatística multivariada, por meio da análise de componentes principais, expressada em forma de gráfico (BIPLOT), utilizando o software R (CORE TEAM, 2015). Esse método foi baseado no método proposto por JOLLIFFE (1972), que seleciona as variáveis próximas aos componentes principais, por estes terem maior correlação com os componentes analisados, que no caso desse estudo as variáveis são as espécies de formigas e os componentes principais são as áreas de estudo.
Na análise geral (área explorada + área não explorada) ocorreram 25 espécies, distribuídas em cinco subfamílias e 17 gêneros. As subfamílias Myrmicinae, Formicinae, Ecitoninae e Ponerinae foram as mais diversificadas na análise geral, com 44%, 24%, 12% e 12% espécies respectivamente. Na área com exploração de madeira, Myrmicinae e Formicinae com 43,48% e 21,74%, respectivamente foram as subfamílias mais representativas em diversidade de espécies, enquanto na área sem exploração de madeira, Myrmicinae e Formicinae com 43,48% e 26,09%, respectivamente, foram as subfamílias mais expressivas (Tabela 1).
Tabela 1. Quantidade de espécies e de indivíduos da subfamília de Formicidae (Hymenoptera)
em cerradão, na área total (A + B), apenas na área com exploração de madeira (A)
e apenas na área sem exploração de madeira (B). Cuiabá, MT. 2011/2012.
Subfamílias |
(A + B) |
|
(A) |
|
(B) |
|||
Espécies |
% |
Espécies |
% |
Espécies |
% |
|||
Ecitoninae |
3 |
12 |
3 |
13,04 |
2 |
8,7 |
||
Formicinae |
6 |
24 |
5 |
21,74 |
6 |
26,09 |
||
Myrmicinae |
11 |
44 |
10 |
43,48 |
10 |
43,48 |
||
Ponerinae |
3 |
12 |
3 |
13,04 |
3 |
13,04 |
||
Pseudomyrmecinae |
2 |
8 |
2 |
8,7 |
2 |
8,7 |
||
Total |
25 |
100 |
23 |
100 |
23 |
100 |
||
Gunawardene et al. (2010) observaram em florestas com e sem corte de madeira uma predominância Myrmicinae, Ponerinae e Formicinae em riqueza de espécies. Souza et al. (2015) amostraram um total de 30 espécies de formigas, sendo 27 em área desmatada e queimada, 19 em vegetação nativa e 18 em área desmatada, sendo a Myrmicinae, a subfamília mais representativa em quantidade de espécies.
A riqueza mensal de espécies de formigas observadas nas duas áreas foi heterogênea, entretanto nos meses de novembro, março e setembro na área com exploração de madeira foram mais expressivas quanto ao número de espécies (Figura 2 A), enquanto que na área sem exploração de madeira, os meses de maio, julho e setembro foram os mais expressivos em quantidade de espécies amostradas (Figura 2 B). Em relação à densidade populacional, na área com exploração de madeira, as maiores quantidades de indivíduos foram coletadas nos meses de Setembro, outubro e novembro, já na área sem exploração de madeira as maiores quantidades de indivíduos ocorreram nos meses de julho, agosto e setembro.
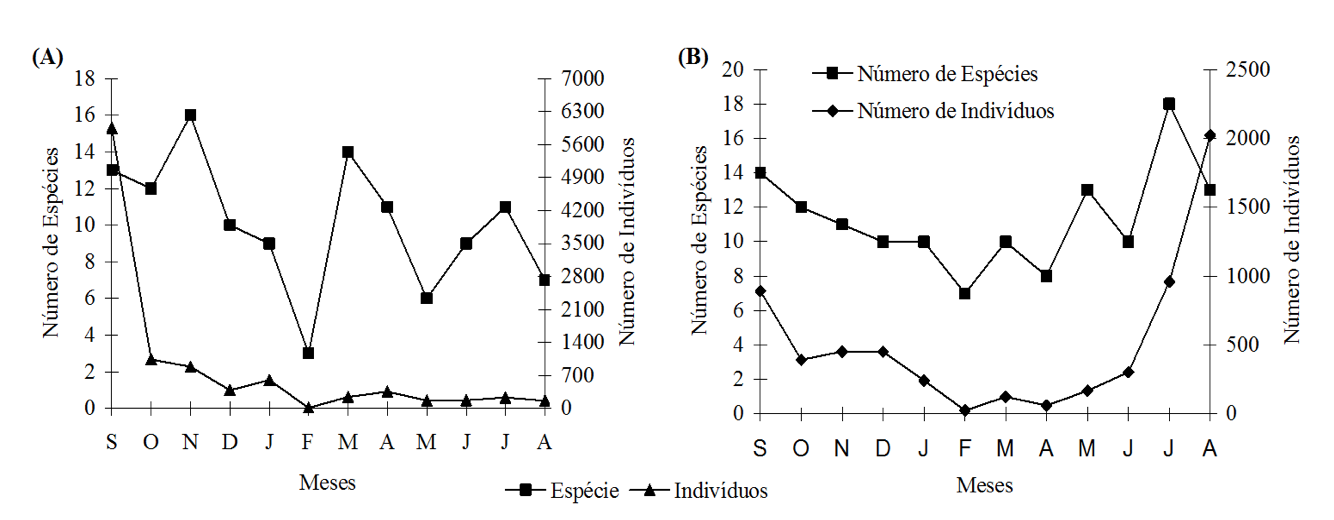
Figura 2. Quantidade mensal de espécies e de indivíduos de Formicidae (Hymenoptera)
em área de vegetação de cerradão com exploração (A) e sem exploração de madeira (B). Cuiabá, MT. 2001.
De acordo com MacKay et al. (1991) áreas abertas dentro de um ambiente florestal ocasionado pela exploração de madeira ou por outras atividades antrópicas, podem propiciar condições ideais para a colonização do local por espécies de formigas generalistas, que são capazes de explorar uma grande diversidade de recursos alimentares e nidificar em diferentes tipos de sítios. Kalif (2001) observou com exceção para as espécies do gênero Pheidole, a exploração de madeira na Amazônia, não afetou a diversidade de espécies de outros gêneros de formigas de serapilheira, mas sim sua composição, com predominância de espécies tolerantes as alterações causadas pela exploração da madeira. Resultados semelhantes foram obtidos por Vasconcelos et al. (2000) e Ketelhut (1999).
Portanto, as espécies com necessidades menos específicas em relação à nidificação e aos hábitos alimentares, tendem a se adaptar com facilidade às alterações ambientais, como a extração de madeiras que, reduz possivelmente as opções de abrigo e alimentos neste ambiente, o que confirma as observações de Brown (1997) de que alterações na diversidade e na composição da entomofauna em florestas tropicais são esperadas após passarem por um distúrbio antrópico.
A ocorrência de alta diversidade de espécies de Myrmicinae confirma que esta é a maior e mais diversificada subfamília de formigas na região Neotropical, caracterizando-se por ser um grupo de formigas abundantes, com facilidade de adaptação a diferentes nichos ecológicos na Região Neotropical, além de várias espécies apresentarem um alto grau de complexidade social (BRANDÃO; CANCELLO, 1999; FERNÁNDEZ, 2003). A subfamília Formicinae é constituída espécies de formigas cosmopolita, apresentando uma tendência na redução do tamanho e ocupação dos microhabitates, tendo várias espécies com hábitos generalistas e predatório não especializado (HOLLDOBLER; WILSON, 1990), podendo colonizar diferentes ambientes, independente de seu estado de preservação, mas da disponibilidade e abundância de alimento para a colônia.
A subfamília Ecitoninae também foi muito expressiva em espécies na área sem extração de madeira, e isto talvez tenha ocorrido em função das espécies desta subfamília ser conhecidas como formiga-de-correição, de hábito de vida nômades, carnívoras e que necessitam de uma grande variedade de hábitats para forragear e devido ao seu hábito alimentar predatório necessitam se deslocar frequentemente a procura de novos recursos alimentares (HOLLDOBLER; WILSON, 1990). Assim, as espécies dessa subfamília podem indicar uma real necessidade de ambientes preservados que, mantenham altos índices de diversidade vegetal e disponibilidade de presas em potencial para suprir o hábito predatório característico das espécies desta subfamília.
Apesar da baixa diversidade de espécies de Ponerinae amostrado, esta subfamília é muito diversificada e amplamente distribuída em todas as regiões tropicais, contendo espécies com hábito alimentar oportunista, predadoras de artrópodes e nectarívoras (CAETANO et al., 2002).
A pouca representatividade de Ponerinae e de Pseudomyrmecinae em ambos os ambientes, pode ter ocorrido em função do local de instalação das armadilhas, das condições ecológicas do local, do hábito alimentar e de nidificação das espécies ou porque naturalmente, sejam espécies que ocorram com baixas densidades populacionais nestas áreas, independente de seu estado de conservação. Porém, Rocha et al. (2015) ao estudarem a diversidade de formigas em Cerrado sensu strictu, coletaram as maiores quantidades de indivíduos de espécies de Ponerinae somente nos ambientes em que a vegetação nativa encontrava-se estabelecida, bem preservado e sem perturbação de origem antrópica, enquanto no ambiente degradado por mineração, não foi constatada a presença de espécies deste gênero de formiga.
O gênero Camponotus destacou-se perante os demais em relação ao número de espécies, com 13,04% no ambiente com exploração e com 17,39% na área sem exploração de madeira (Tabela 2).
Tabela 2. Quantidade de Espécies (E) da mirmecofauna coletada nas armadilhas
em área com extração de madeira (A) e sem extração de madeira (B) no município de Cuiabá.
Subfamília/Gêneros |
(A) |
(B) |
||
E |
% |
E |
% |
|
Ecitoninae |
|
|
|
|
Labidus |
2 |
8,70 |
2 |
8,70 |
Nomamyrmex |
1 |
4,35 |
- |
- |
Formicinae |
|
|
|
|
Brachymyrmex |
1 |
4,35 |
1 |
4,35 |
Camponotus |
3 |
13,04 |
4 |
17,39 |
Nylanderia |
1 |
4,35 |
1 |
4,35 |
Myrmicinae |
|
|
|
|
Atta |
1 |
4,35 |
1 |
4,35 |
Basiceros |
1 |
4,35 |
1 |
4,35 |
Cephalotes |
2 |
8,70 |
2 |
8,70 |
Crematogaster |
1 |
4,35 |
1 |
4,35 |
Pheidole |
2 |
8,70 |
2 |
8,70 |
Solenopsis |
1 |
4,35 |
1 |
4,35 |
Strumigenys |
- |
- |
1 |
4,35 |
Trachymyrmex |
2 |
8,70 |
1 |
4,35 |
Ponerinae |
|
|
|
|
Gnamptogenys |
1 |
4,35 |
1 |
4,35 |
Pachycondyla |
2 |
8,70 |
2 |
8,70 |
Pseudomyrmecinae |
|
|
|
|
Odontomachus |
1 |
4,35 |
1 |
4,35 |
Pseudomyrmex |
1 |
4,35 |
1 |
4,35 |
Total |
23 |
100 |
23 |
100 |
De acordo com Silvestre et al. (2003) as espécies dos gêneros Labidus e Pheidole caracterizam-se por populações oportunistas e generalistas em relação à nidificação, pois constroem seus ninhos em locais como galhos e troncos podres de árvores, no solo, em montes de matéria orgânica, embaixo de rochas e associado à raízes de plantas, nidificando preferencialmente em troncos podres caídos no chão da floresta e algumas espécies constroem ninhos de palha seca e gravetos.
O gênero Solenopsis abrange espécies cosmopolitas, conhecidas por "formigas-de-fogo" com ninhos subterrâneos e caracterizando-se pela onivoria. Segundo Bueno & Campos-Farinha (1998), as operárias das espécies deste gênero são bastante agressivas e ativas na procura de alimentos, podendo colonizar diferentes habitats. De acordo com Fowler et al. (1991) e Delabie e Fowler (1995) as espécies do gênero Solenopsis são agressivas na utilização dos recursos ao nível da serapilheira, suportando longos períodos com baixa oferta de alimento, além de apresentarem estratégias de recrutamento muito eficiente.
As espécies do gênero Camponotus caracterizam-se por ser o mais diversificado em espécies da região Neotropical, explorando uma grande variedade de ambientes e seus ninhos, geralmente são encontrados no solo, em ramos, galhos podres e raramente em madeira viva (BOLTON, 1973; 1994) e a maioria das espécies são onívoras, podendo se alimentar de secreções de plantas, honeydew de hemípteros sugadores de seivas (RICO-GRAY; OLIVEIRA, 2007).
Observa-se que em ambas as áreas a riqueza de espécies amostradas foram semalhantes. Miranda et al. (2013) constataram que dependendo da intensidade de manejo utilizado em extração de madeira em empreendimento agroflorestal, pode não alterar de modo significativo à riqueza de formigas no local. Corroborando em parte com estas afirmações, Vanderwoude e Lobry de Bruyn (2000) observaram em área com plantio de Corymbia citriodora submetida ao manejo florestal, pequenas diferenças na composição da comunidade de formigas entre as áreas com e sem corte de árvores e afirmaram que o resultado observado deve-se ao modelo de manejo florestal adotado.
Azevedo-Ramos et al. (2006) detectaram um aumento na riqueza de formigas após o manejo florestal de baixo impacto em floresta nativa, atribuindo aumento à intensidade de extração de madeira mais elevada utilizada, indicando que dependendo do grau de intensidade do manejo e o tempo de pós-exploração podem influenciar diretamente a diversidade de espécies de formigas dentro de áreas, mesmo sob manejo adequado.
Estudos de Woodcock et al. (2013) constataram que o efeito causado pela extração seletiva de madeira intensiva em floresta tropical foi alteração nos níveis tróficos de 159 espécies de formigas de serapilheira e uma diminuição de 30% na abundância de formigas na área sob exploração de madeira. Dittmann et al. (2014) observaram que em floresta sob regime de exploração seletiva de madeira, a densidade, o tamanho das colônias e o hábito alimentar das espécies de formigas foram diferentes quando comparados com colônias em uma floresta primária intacta e em uma área exposta à perturbação natural causado por um rio adjacente.
O trabalho de Gunawardene et al. (2010) mostrou que embora não tenha ocorrido uma diferença significativa na riqueza de espécies e abundâncias de formigas entre a floresta explorada e não explorada, pode-se observar que o corte seletivo tem impacto mínimo sobre a riqueza de espécies de formigas em uma determinada floresta, independentemente da intensidade de exploração madeireira, pois, após um determinado tempo, a floresta pode recuperar sua riqueza de espécies de formigas observada antes da execução do manejo florestal.
3.1. Identificação das espécies de formigas coletadas nos ambientes
Do total ocorreram 25 espécies amostradas, sendo 21 comuns aos dois ambientes e três restrita para cada área amostrada, como as espécies Nomamyrmex esenbecki (Ecitoninae), Trachymyrmex urichi (Myrmicinae) que foram restritas a área com exploração, enquanto Camponotus sp. (Formicinae) e Strumigenys sp. (Myrmicinae) ocorreram exclusivamente na área sem exploração de madeira (Tabela 3). Na área com exploração de madeira, S. invicta, P. rodeszkowii, A. sexdens rubropilosa e C. rufipes com 53,56%, 20,28%, 11,96% e 7,25%, respectivamente, foram as espécies com as maiores quantidades de indivíduos coletados, enquanto na área sem exploração, L. praedator, A. sexdens rubropilosa e P. radoszkowskii, com 39,26%, 19,96% e 19,81%, respectivamente, foram as espécies mais abundantes em espécimes coletados.
Tabela 3. Quantidade de indivíduos (I) das espécies formigas identificadas nas armadilhas coletadas
na área com extração de madeira (A) e sem extração de madeira (B) no município de Cuiabá.
Subfamília/Espécie |
Área A |
Área B |
Total |
||
I |
% |
I |
% |
||
Ecitoninae |
|
|
|
|
|
Labidus praedator (Fr. Smith, 1858) |
166 |
1,62 |
2.400 |
39,26 |
2.566 |
Labidus spininodis Emery, 1890 |
181 |
1,76 |
108 |
1,77 |
289 |
Nomamyrmex esenbecki Westwood, 1842 |
1 |
0,01 |
- |
- |
1 |
Formicinae |
|
|
|
|
|
Brachymyrmex pilipes Mayr, 1887 |
11 |
0,11 |
108 |
1,77 |
119 |
Camponotus cingulatus Mayr, 1862 |
39 |
0,38 |
3 |
0,05 |
42 |
Camponotus crassus Mayr, 1862 |
18 |
0,18 |
56 |
0,92 |
74 |
Camponotus rufipes (Fabricius, 1775) |
744 |
7,25 |
181 |
2,96 |
925 |
Camponotus sp. |
- |
- |
1 |
0,02 |
1 |
Nylanderia sp. |
92 |
0,90 |
151 |
2,47 |
243 |
Myrmicinae |
|
|
|
|
|
Atta sexdens rubropilosa Forel, 1908 |
1.227 |
11,96 |
1.220 |
19,96 |
2.447 |
Basiceros balzani (Emery, 1894) |
20 |
0,19 |
5 |
0,08 |
25 |
Cephalotes atratus (Linnaeus, 1758) |
1 |
0,01 |
4 |
0,07 |
5 |
Cephalotes pusillus (Klug, 1824) |
2 |
0,02 |
1 |
0,02 |
3 |
Crematogaster stollii Forel, 1885 |
11 |
0,11 |
24 |
0,39 |
35 |
Pheidole radoszkowskii Mayr, 1884 |
2.081 |
20,28 |
1.211 |
19,81 |
3.292 |
Pheidole fimbriata Roger 1863 |
8 |
0,08 |
198 |
3,24 |
206 |
Solenopsis invicta Buren, 1972 |
5.495 |
53,56 |
249 |
4,07 |
5.744 |
Strumigenys sp. |
- |
- |
5 |
0,08 |
5 |
Trachymyrmex urichi Forel, 1893 |
4 |
0,04 |
- |
- |
4 |
Trachymyrmex sp. |
2 |
0,02 |
12 |
0,20 |
14 |
Ponerinae |
|
|
|
|
|
Gnamptogenys moelleri (Forel, 1912) |
18 |
0,18 |
16 |
0,26 |
34 |
|
|
|
|
Continua... |
|
Continuação da Tabela 3 |
|
|
|
|
|
Subfamília/Espécie |
Área A |
Área B |
Total |
||
I |
% |
I |
% |
||
Pachycondyla verenae (Forel, 1922) |
39 |
0,38 |
82 |
1,34 |
121 |
Pachycondyla sp. |
80 |
0,78 |
23 |
0,38 |
103 |
Pseudomyrmecinae |
|
|
|
|
|
Odontomachus bauri Emery, 1891 |
8 |
0,08 |
27 |
0,44 |
35 |
Pseudomyrmex tenuis (Fabricius, 1804) |
12 |
0,12 |
28 |
0,46 |
40 |
Total |
10.260 |
100 |
6.113 |
100 |
16.373 |
I= Indivíduos
De acordo com Hölldobler e Wilson (1990), espécies do gênero Solenopsis apresentam características agressivas e são bioindicadores de ambientes alterados. Estudos de Almeida et al. (2007) relataram que S. invicta apresenta maiores quantidades de indivíduos em ambientes mais simplificados (monocultivos).
A ocorrência de L. praedator na área sem exploração de madeira justifica-se, pois é uma espécie de formiga-de-correição, carnívoras, bastante agressivas e geralmente exploram e coloniza diferentes hábitats a procura de alimento ou de presas para a manutenção de sua colônia. Segundo Brandão et al. (2000), L. praedator é uma espécie epigéica, nômade e predadora de outros artrópodes e são de hábitos quase que exclusivamente subterrâneos, com inúmeros orifícios de entrada na superfície do solo e podem ser encontradas em grandes quantidades na serapilheira.
A importância da característica, composição e espessura serapilheira representa maior oferta de alimento e locais para nidificação, pois tem um microclima favorável à vida de várias espécies de formigas (NAKAMURA et al., 2003). Portanto, um dos fatores que pode determinar a abundância de indivíduos de várias espécies de formigas dentro de um determinado ambiente florestal é a quantidade de presas em potencial que, normalmente são mais abundantes na serapilheira e que são, normalmente, mais expressivas em áreas não antropizadas. Contudo, os resultados obtidos neste trabalho não corroboram com as afirmações de Matos et al. (1994) e de Oliveira et al. (1995) que condicionaram o aumento na diversidade de espécies de formigas com o grau de complexidade da vegetação e da serapilheira e que a variação na riqueza de espécies de formigas é influenciada pelas características do ambiente, pois quanto maior for sua complexidade, maior será a diversidade de espécies de formicídeos no local.
3.2. Índice de Diversidade dos ambientes amostrados
Na Tabela 4, pode-se observar que a riqueza de espécies nos dois ambientes amostrados foi igual, porém com diferenças na sua composição. Contudo, na área sem exploração de madeira, ocorreu um maior número de espécies abundantes (N1= 6,20) e muito abundantes (N2= 4,18), indicando que uma maior quantidade de espécies conseguiu explorar com maior eficiência os recursos ambientais disponíveis para sua reprodução, manutenção e colonização da área, evitando-se assim a dominância de poucas espécies.
Tabela 4. Índice de Diversidade de acordo com a série de números de Hill e Equitatividade das espécies
de formigas amostradas em vegetação de cerrado com e sem extração de madeira.
ÁREA |
N0 |
N1 |
N2 |
H’ |
ʎ |
E |
Área com extração de madeira |
23 |
4,21 |
2,87 |
1,43 |
0,34 |
0,58 |
Área sem extração de madeira |
23 |
6,20 |
4,18 |
1,82 |
0,23 |
0,61 |
(N0)= número total de espécies; (N1) = número de espécies abundantes;
(N2) = número de espécies muito abundantes; (H’) = índice de Shannon Winner;
(ʎ) = índice de Simpson; (E) = equitatividade.
Os componentes básicos da diversidade de Shannon-Winner, riqueza e equitatibilidade, apresentaram resultados diferentes, pois enquanto a riqueza foi igual em ambas as áreas, a equitatibilidade, que mede a distribuição dos indivíduos dentro das populações, foi muito diferente nos dois ambientes, sendo mais homogêneo na área sem exploração de madeira, indicando que o local contém recursos alimentares e ecológicos em quantidade e qualidade suficiente para que maior número de espécies de formigas ocorresse com densidades populacionais sem grandes diferenças entre si, evitando assim, a dominância de poucas espécies de formigas na exploração dos recursos disponíveis.
Um fator que pode ter influenciado na densidade populacional das espécies de formigas nas duas áreas, foram as chuvas constantes na região, entre os meses de novembro e maio, causando acúmulo de água na superfície do solo, que pode ter dificultado ou mesmo impedido o acesso e a colonização por espécies de formigas oportunistas com grandes densidades populacionais, principalmente na área sem exploração de madeira, pois devido á cobertura florestal inalterada, esta área permaneceu inundada por um maior período de dias, permanecendo no local com densidades populacionais mais homogêneas, apenas as espécies de formigas nativas já adaptadas a esta alteração ambiental temporária. Linksvayer e Janssen (2009) afirmaram que o alagamento favorece as espécies de formigas oportunistas, pois são mais adaptadas em colonizar áreas alteradas ou antropizadas e caracterizando-se pelo pequeno tamanho da colônia e dieta não especializada.
O local do experimento caracteriza-se por período seco muito rigoroso, com altas temperaturas e baixa umidade relativa o que afeta profundamente as condições ambientais, principalmente junto à superfície do solo, devido maioria das espécies vegetais perderem parte de suas folhas neste período, permitindo uma maior penetração dos raios solares, tornando o local inadequado para as espécies de formigas que necessitam de um ambiente com condições mais favoráveis para colonizar.
A precipitação pluvial pode ter influenciado na sazonalidade, afetando as densidades populacionais de algumas espécies de formigas no local, devido ao tipo de solo da região, a água da chuva não penetra profundamente no solo, formando grandes áreas alagadas por vários dias, afetando as condições da serapilheira, inviabilizando o deslocamento para a procura de alimento e construção de novos ninhos. Mertl et al. (2009) observaram em floresta tropical, que as inundações alteram de forma significativa a riqueza e composição das assembleias de formigas de serapilheira, pois causam uma diminuição na riqueza de espécies e na quantidade de ninhos e Ballinger et al. (2007) também constataram uma diminuição na riqueza de formigas após uma inundação em uma área de floresta e que o impacto mais significativo ocorreu nas áreas que permaneceram alagadas por um maior período de tempo.
O gráfico Biplot representa simultaneamente as coordenadas com ocorrência das espécies de formigas em função dos locais amostrados para o período de estiagem ou chuvoso, a soma da variabilidade retida nos componentes explicou 100% da variabilidade original, onde CP1 e CP2 retêm cada um, 53,57% e 46,43% respectivamente, das informações originais dos dados obtidos no período de chuvas (Figura 3), assim é possível afirmar que algumas espécies de formigas apresentam ocorrência influenciada em função do ambiente amostrado, pois pode ser observado que as espécies: 5- S. invicta apresenta maior associação com o ambiente explorado, já as espécies 3- P. radoszkowskii e 12- L. praedador com o ambiente sem exploração.
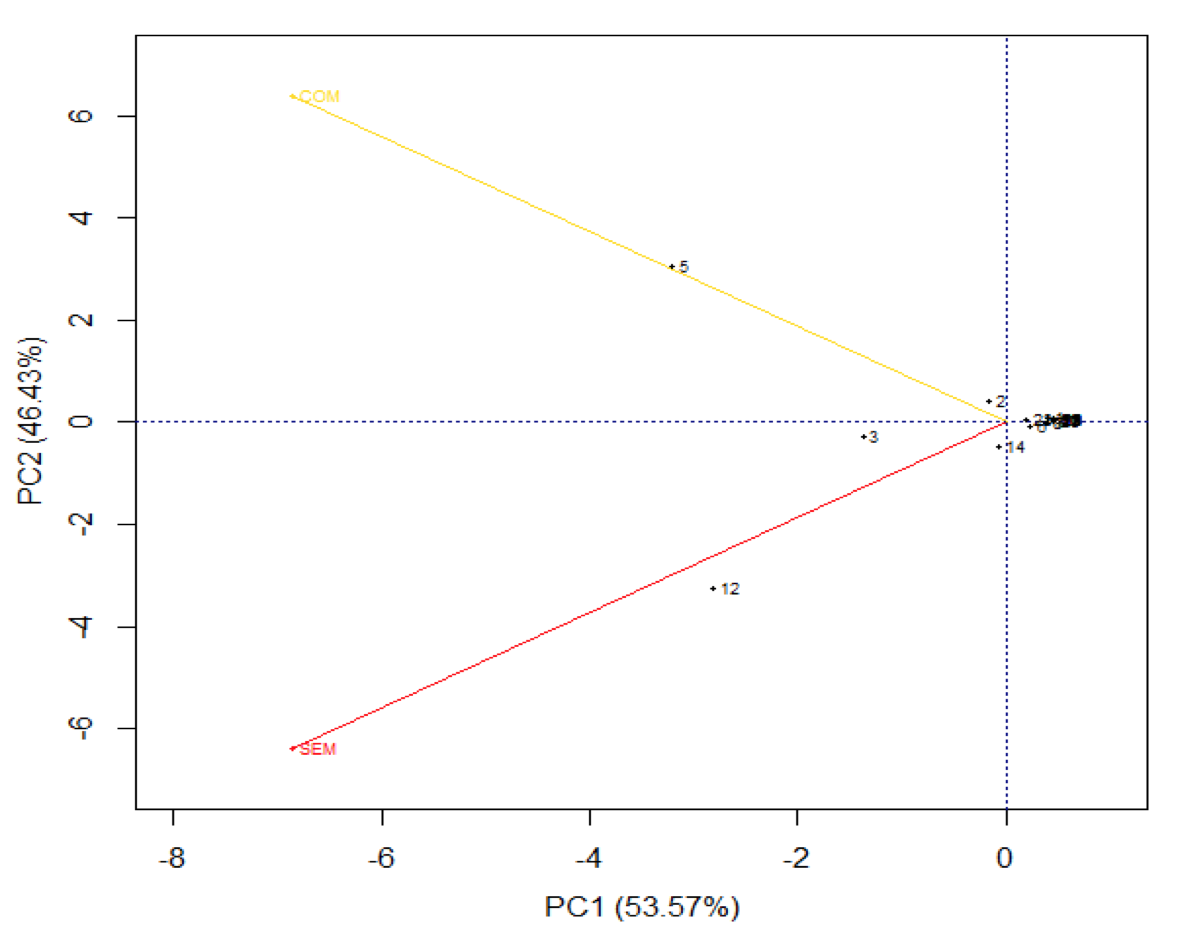
Figura 3. Análise de componentes principais utilizando gráfico BIPLOT com ocorrência
das espécies de formigas em função dos locais amostrados para o período de chuvas.
No período de estiagem a soma da variabilidade retida nos componentes explicou 100% da variabilidade original, onde CP1 e CP2 retêm cada um, 88,22% e 11,78% respectivamente (Figura 4), assim é possível afirmar que algumas espécies de formigas apresentam ocorrência influenciada em função do ambiente amostrado, pois pode ser observado que as espécies: 5- S. invicta e 3- P. radoszkowskii apresentam maior associação com o ambiente explorado, já a espécie 14- A. s. rubropilosa com o ambiente sem exploração.
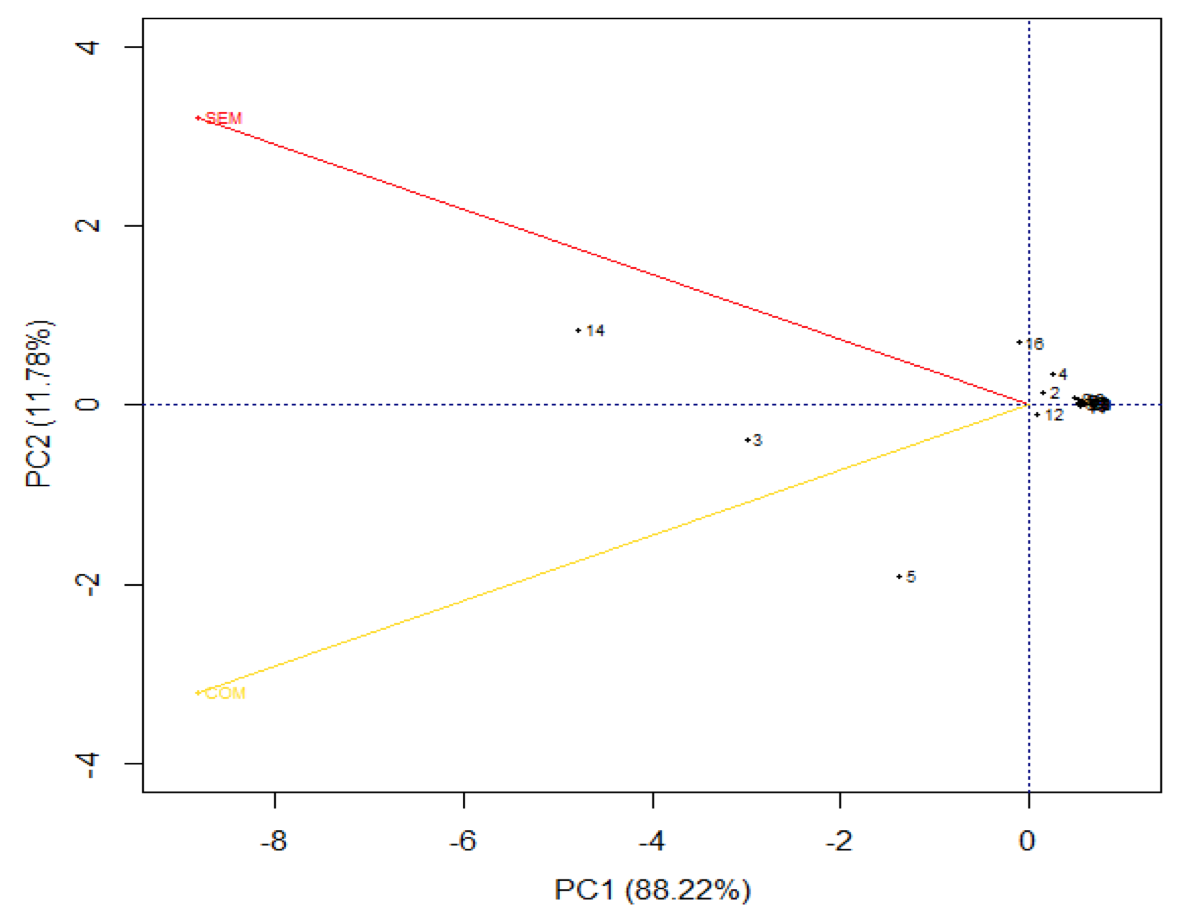
Figura 4. Análise de componentes principais utilizando gráfico BIPLOT com ocorrência das espécies
de formigas em função dos locais amostrados para o período de estiagem.
Pode se perceber que P. rodeszkowskii é influenciada pelas mudanças do ambiente, pois na época de chuva a espécie teve associação com o ambiente sem exploração, mudando de comportamento na época de estiagem, associando com o ambiente explorado. De fato a espécie é dominante em ambientes perturbados (PERFECTO, 1994). De acordo com Perfecto e Vandermeer (1996), P. rodeszkowskii é ideal para examinar feitos diretos e indiretos das mudanças de microhábitat na diversidade de espécies, devido à adaptabilidade desta sobre ambientes, principalmente ambientes explorados, uma vez que a espécie se sobressai em relação aos demais formicídeos. Assim essa mudança de comportamento pode estar associada a ocorrência das demais espécies presentes no ambiente em função do período climático.
Outra questão pode ser observada com a espécie S. invicta, que independente do período climático, teve associação com o ambiente explorado, sendo essa espécie comum em ambientes perturbados (PERFECTO,1994). Já associação da espécie A. s. rubropilosa com ambiente natural na época de seca pode ter relação à escassez de espécies vegetais no ambiente de cerrado, ainda mais quando há a exploração de madeira, fazendo com que as formigas cortadeiras forrageiem locais com maior diversidade de plantas.
- A riqueza de espécies de formigas não é afetada pela exploração de madeira em função da rápida recuperação das condições ambientais que ocorre anualmente na vegetação de cerrado;
- As densidades populacionais das espécies de hábito alimentar generalista e oportunista foram mais abundantes em número de indivíduos, independente das condições das áreas amostradas;
Ao professor Dr. Jacques Hubert Charles Delabie, da CEPLAC, de Ilhéus/BA, pela identificação taxonômica das espécies e ao Técnico Manoel Lauro da Silva, do Laboratório de Proteção Florestal (LAPROFLOR/FENF/UFMT) pelo inestimável auxílio na realização deste trabalho.
ALMEIDA, F. S.; QUEIROZ, J. M.; MAYHÉ-NUNES, A. J. (2007). Distribuição e abundância de ninhos de Solenopsis invicta Buren (Hymenoptera: Formicidae) em um agroecossistema diversificado sob manejo orgânico. Floresta e Ambiente, 14(1), 33-43.
ALONSO, L. E. & DONATI, A. (2000). Biodiversity studies, monitoring and ants: an overview, p. 1-8. In: AGOSTI, D.; MAJER, J.D.; ALONSO, L.E.; SCHULTZ, T.R.(editores). Ants: standard methods for measuring and monitoring biodiversity. Washington: Smithsonian Institution Press,
AZEVEDO-RAMOS, C.; CARVALHO JUNIOR, O.; AMARAL, B. D. (2006). Short-term effects of reduced impact logging on eastern Amazon fauna. Forest Ecology and Management, 232(1), 26-35.
BALLINGER, A.; LAKE, P. S.; MAC-NALLY, R. (2007) Do terrestrial invertebrates experiences floodplains as landscape mosaics? Immediate and longer-term effects of flooding on ant assemblages in a floodplain forest. Oecologia, 152(1), 227-238.
BOLTON B. (1973). The ants genera of West Africa: a synonymic synopsis with keys (Hymenoptera: Formicidae). Bulletin of the Britsh Museum Entomology, 27(1), 317-368.
BOLTON, B. (1994). Identification guide to ant genera of the world. Cambridge: Harvard University Press, 222p.
BUENO, O. C.; CAMPOS-FARINHA, A. E. C. (1998). Formigas Urbanas: Comportamento das espécies que invadem as cidades brasileiras. Vetores & Pragas, 12(5), 13–16.
BRANDÃO, C. R. F.; CANCELLO, E. M. (1999). Invertebrados Terrestres. Vol. V Biodiversidade do Estado de São Paulo. Síntese do conhecimento ao final do século XX. JOLY, C. A.; BICUDO, C. E. M. (orgs). São Paulo, FAPESP. 279p.
BRANDÃO, C. R. F.; CANCELLO, E. M.; YAMAMOTO, C. I. (2002). Invertebrados terrestres. In: Lewinsohn TM (org.). Avaliação do estado do conhecimento da biodiversidade brasileira: Sumário Executivo. Brasília: MMA.
BROWN, K. S. (1997). Diversity, disturbance and sustainable use of Neotropical forests: insects as indicators for conservation monitoring. Journal Insect Conservation, 1(1), 25-42.
CAETANO, F. H.; JAFFÉ, K.; ZARA, F. J. (2002). Formigas: Biologia e anatomia. São Paulo: Ed. Rio Claro; 131p.
CAMPOS, R. I.; LOPES, C. T.; MAGALHÃES, W. C. S.; VASCONCELOS, H. L. (2008). Estratificação vertical de formigas em Cerrado strictu sensu no Parque Estadual da Serra de Caldas Novas, Goiás, Brasil. Iheringia, Série Zoológica, 98(3), 311-316.
CORE TEAM. R. (2016); A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Disponível em: <URL https://www.R-project.org/> Acessado em 19/09/2016.
CORRÊA, M. M.; FERNANDES, W. D.; LEAL, I. R. (2006). Diversidade de formigas epigéicas (Hymenoptera: Formicidae) em capões do Pantanal Sul Matogrossense: Relações entre riqueza de espécies e complexidade estrutural da área. Neotropical Entomology, 35(6), 724-730.
DELABIE, J. H. C.; FOWLER, H. G. (1995). Soil and litter cryptic ant assemblages of Bahian cocoa plantations. Pedobiologia, 39(5), 423-433.
DITTMANN, M. T,; DAMMHAHN, M.; KAPPELER, P. M. (2014). Investigating behavior and ecology of Aphaenogaster swammerdami (Formicidae) in selectively logged forest: 20 years later – a happy ant?. Malagasy Nature, 8(1), 35-48.
FERNÁNDEZ, F. (2003). Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de Investigacion de Recursos Biológicos Alexander von Humboldt, 398p.
PHILPOTT S. M.; PERFECTO, I.; ARMBRECHT, I.; PARR, C. L. (2010). Ant diversity and function in disturbed and changing habitats, In LACH, L., PARR, C. L., ABBOTT, K. L. (Eds), Ant Ecology (pp.137–156). Oxford University Press, Oxford, UK. 194p.
FREIRE, C. B.; OLIVEIRA, G. V.; MARTINS, F. R. S.; SOUZA, L. E. C.; RAMOS-LACAU, L. S.; CORRÊA, M. M. (2012). Riqueza de formigas em áreas preservadas e em regeneração de caatinga arbustiva no sudoeste da Bahia, Brasil. Revista Brasileira de Biociências, 10(1), 131-134.
FREITAS, A. V. L.; FRANCINI, R. B.; BROWN Jr., K. S. (2004). Insetos como indicadores ambientais. In CULLEN Jr., L.; RUDRAN, R.; VALLADARES-PADUA, C. (Org.). (pp. 125-151). Métodos de estudos em biologia da conservação e manejo da vida silvestre. Editora UFPR, Curitiba. 665p.
FOWLER, H. G. L.; FORTI, C.; BRANDÃO, C. R. F.; DELABIE, J. H. C.; VASCONCELOS, H. L. (1991). Ecologia nutricional de formigas. In PAZZINI, R. & PARRA, J. R. P. (eds). Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo, Manole, p.131-209.
GUNAWARDENE, A. N. R.; MAJER, J. D.; EDIRISINGHE, A. J. P. (2010). Investigating residual effects of selective logging on ant species assemblages in Sinharaja Forest Reserve, Sri Lanka. Forest Ecology and Management, 259(1), 555–562.
HOLLDOBLER, B.; WILSON, E. O. 1990. The ants. Cambridge: Havard University Press, 746p.
JOLLIFFE, I. T. (1972); Discarding Variables in a Principal Component Analysis. I: Artificial Data. Journal of the Royal Statistical Society. Series C (Applied Statistics), 21(2), 160-173.
KALIF, K. A. B.; MOUTINHO, P. R. S. (2000). Compararison of tree methods of sampling ant in a tropical forest on area in eastern Amazônia. Boletim of Museu Paraense Emílio Goeldi. Série Zoológica, 16(1), 75-81.
KETELHUT, S. M. (1999). Avaliação das comunidades de formigas em uma área de extração de madeira. Dissertação (Mestrado em Zoologia) - Universidade Federal do Pará e Museu Paraense Emilio Goeldi. 110p.
LINKSVAYER, T. A.; JANSSEN, M. A. (2009). Traits underlying the capacity of ant colonies to adapt to disturbance and stress regimes. Systems Research and Behavioral Science, 26(3) 315-329.
LOBRY DE BRUYN L. A. (1999). Ants as bioindicators of soil function in rural environments. Agriculture, Ecosystems & Environment, 74(1), 425-441.
MAJER, J. D. (1983). Ants: bioindicadores of mine site rehabilitation, land use and land conservation. Environmental Management, 7(4) 375-383.
MAJER, J. D. (1992). Ant recolonization of rehabilitated bauxite mines of Poços de Caldas, Brasil. Journal of Applied Ecology, 8(1), 97-108.
MAJER J.D. (1996). Ant recolonization of rehabilitated bauxite mines at Trombetas, Pará, Brazil. Journal of Applied Ecology, 12(2), 257-273.
MAJER, J. D.; NICHOLS, O. G. (1998). Long-term recolonization patterns of ants in Western Australian rehabilitated bauxite mines with reference to their use as indicators of restoration success. Journal of Applied Ecology, 35(1), 161-182.
MATOS, J. A.; YAMANAKA, C. N.; CASTELLANI, T. T.; LOPES, B. C. (1994). Comparação da fauna de formigas de solo em áreas de plantio de Pinus elliottii, com diferentes graus de complexibilidade estrutural (Florianópolis, SC.). Biotemas, 7(1/2), 57-64.
MACKAY, W. P.; REBELES, A. M.; ARREDONDO, H. C. B.; RODRIGUEZ, A. D. R.; GONZALEZ , D. A.; VINSON, S. B. (1991). Impact of the slashing and burning of a tropical rain forest on the native ant fauna (Hymenoptera: Formicidae). Sociobiology, 18(3), 257-268.
MERTL, A. L.; RYDER, W.K.T.;TRANIELLO, J. F. A. (2009). Impact of flooding on the species ricchness, density and composition of Amazonian litter-nesting ants. Biotrópica, 41(5), 633-641.
MIRANDA, P. N.; MORATO, E. F.; OLIVEIRA, M. A.; DELABIE, J. H. C. (2013). A riqueza e composição de formigas como indicadores dos efeitos do manejo florestal de baixo impacto em floresta tropical no estado do Acre. Revista Árvore, 37(1), 163-173.
NAKAMURA, A.; PROCTOR, H.; CATTERALL, C. P. (2003). Using soil and litter arthropods to assess the state of rainforest restoration. Ecological Management & Restoration, 4(1), 20-28.
OLIVEIRA, M. A.; DELLA LUCIA, T. M. C.; ARAÚJO, M. S; CRUZ, A. P. (1995). A fauna de formigas em povoamentos de eucalipto na mata nativa no estado do Amapá. Acta Amazônica, 25(1/2), 117-126.
OLSON, D.M. (1991). A comparison of the efficacy of litter sifting and pitfall traps for sampling leaf litter ants (Hymenoptera, Formicidae) in a Tropical Wet Forest, Costa Rica. Biotropica, 23(2), 166-172.
PERFECTO. I.; VANDERMEER, J. (1996) Microclimatic changes and the indirect loss of ant diversity in a tropical agroecosystem. Oecologia, 108(3), 577-582.
PERFECTO. I. (1994) Foraging behavior as a determinant of asymmetric competitive interaction between two ant species in a tropical agroecosystem. Oecologia, 98(2), 184-192.
RICO-GRAY, V. & OLIVEIRA, P. S. (2007). The ecology and evolution of ant-plant interactions. Chicago: University of Chicago Press, 320 p.
ROCHA, W. O.; DORVAL, A.; PERES FILHO, O.; VAEZ, C. A.; RIBEIRO, E. S. (2015). Formigas (Hymenoptera: Formicidae) bioindicadoras de degradação ambiental em Poxoréu, Mato Grosso, Brasil. Floresta e Ambiente, 22(1), 88-98.
SANTOS, M. S.; LOUZADA, J. N. C.; DIAS, N. (2006). Litter ants richness (Hymenoptera: Formicidae) in remnants of a semi-dicidous forest in the Atlantic rain forest, Alto do Rio Grande region, Minas Gerais. Ihleringia, série Zoologia, 96(1), 95-101.
SILVA, R. R,; BRANDÃO, C. R. F. (1999). Formigas (Hymenoptera: Formicidae) como indicadores da qualidade ambiental e da biodiversidade de outros invertebrados terrestres. Biotemas, 12(2), 55-73.
SILVESTRE, R.; BRANDÃO, C. R. F.; DA SILVA, R. R. (2003). Grupo funcionales de hormigas: el caso de los grêmios del Cerrado. In: FERNÁNDEZ F. (Editor). Introducción a las hormigas de La región Neotropical.Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, p.113-148.
SOARES, S. A.; ANTONIALLI-JÚNIOR, W. F.; LIMA-JÚNIOR, S. E. (2010). Diversidade de formigas epigéias (Hymenoptera, Formicidae) em dois ambientes no Centro-Oeste do Brasil. Revista Brasileira de Entomologia, 54(1), 76-81.
SOUSA, I. D.; MARINHO, C. G. S.; LIMA, A. S.; MELO, B. A.; OLIVEIRA, M. A.; DELABIE, J. H. C. (2015). Diversidade de formigas (Hymenoptera: Formicidae) epigéias na mesoregião do sertão paraibano. Revista Brasileira de Zoociências, 16(1), 43-53.
SUGUITURU, S. S.; SOUZA, D. R.; MUNHAE, C. B, PACHECO, R.; MORINI, M. S. C. (2013). Diversidade e riqueza de formigas (Hymenoptera: Formicidae) em remanescentes de Mata Atlântica na Bacia Hidrográfica do Alto Tietê, SP. Biota Neotropica, 13(2), 141-152.
VANDERWOUDE, C.; LOBRY DE BRUYN, L. A. (2000). Short-term responses by ants (Hymenoptera: Formicidae) to selective timber harvesting of an open forest in eastern Australia. Australian Forestry, 63(4), 267-273.
VASCONCELOS, H. L.; VILHENA, J. M. S.; CALIRI, G. J. A. (2000). Responses of ants to selective logging of a central Amazonian forest. Journal of Applied Ecology, 37(1), 1-8.
WILSON, E. O. (1971). The insect societies. Harvard University press, Cambrigde, Mass, 562p.
WOODCOCK, P.; EDWARDS, D. P.; NEWTON , R. J.; KHEN,C. V.; BOTTRELL ,S. H.; HAMER, K. C. (2013). Impacts of Intensive Logging on the Trophic Organisation of Ant Communities in a Biodiversity Hotspot. Journal Plos One, 8(4), 1-7.
1. Professor Doutor da Faculdade de Engenharia Florestal da Universidade Federal de Mato Grosso (Cuiabá), como Professor Adjunto (DE) - Área de Ecologia de insetos.
2. Professor Doutor da Faculdade de Engenharia Florestal da Universidade Federal de Mato Grosso (Cuiabá), como Professor Adjunto (DE) - Área de entomologia florestal.
3. Biólogo e Mestre em Ciências florestais e Ambientais – Área de Entomologia.
4. Professor Doutor da Faculdade de Engenharia Ambiental da Universidade de Cuiabá, como Professor Celetista (CLT) – Área de Estatística.
5. Professor Mestre da Faculdade de Engenharia Ambiental do Centro Universitário de Várzea Grande, Universidade de Cuiabá, como Professor Celetista (CLT) – Área de avaliação de impactos ambientais.