![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 23) Año 2017. Pág. 26
Leonardo Julian SULZBACHER 1; Valvenarg Pereira da SILVA 2; Bruno Wagner ZAGO 3; Carla Lima CORRÊA 4; Aline Vidor Melão DUARTE 5; Marco Antonio Aparecido BARELLI 6
Recibido: 06/12/2016 • Aprobado: 25/01/2017
RESUMO: Esta pesquisa teve como objetivo avaliar a divergência genética entre 21 genótipos de feijão e sugerir combinações híbridas superiores. O delineamento experimental utilizado foi de blocos ao acaso com três repetições. Os dados foram submetidos a análise de variância, teste de Scott e Knott, métodos aglomerativos de otimização de Tocher e Hierárquico UPGMA e variáveis canônicas, fundamentados na distância generalizada de Mahalanobis. Genótipos superiores podem ser obtidos a partir das hibridações: BRS Majestoso x Iraí, BRS Majestoso x Jalo Precoce, BRS Majestoso x BRS Radiante, BRS Grafite x Iraí, VC 6 x Jalo Precoce e BRS Horizonte x BRS Radiante. |
ABSTRACT: This study aimed to evaluate the genetic divergence between 21 common bean genotypes and suggest superior hybrid combinations. The experimental design was a randomized block with three replications. Data were subjected to analysis of variance, Scott and Knott cluster analysis, the agglomerative optimization Tocher’s method and hierarchical UPGMA as well as canonical variables analysis, based on the Mahalanobis distance. Superior genotypes can be obtained by hybridization of the following genotypes: BRS Majestoso x Iraí, BRS Majestoso x Jalo Precoce, BRS Majestoso x BRS Radiante, BRS Grafite x Iraí, VC 6 x Jalo Precoce e BRS Horizonte x BRS Radiante |

O feijão comum (Phaseolus vulgaris L.) é produzido em praticamente todo o território brasileiro, constitui-se como uma das principais fontes de proteínas e de aminoácidos essências na dieta da população brasileira (Tsutsumi et al., 2015). Apresenta uma ampla variabilidade genética quanto aos tipos cultivados, sendo estes pertencentes aos grupos de origem mesoamericano (sementes pequenas) e andino (sementes grandes). Esses grupos têm sido subdivididos dentro de raças, baseados principalmente em avaliações morfológicas, agronômicas e bioquímicas (Singh et al., 1991).
Segundo Rocha et al. (2014), a existência desta variabilidade genética dentro e entre estes grupos é fundamental para o sucesso em programas de melhoramento de plantas autógamas, tendo em vista que o conhecimento desta variabilidade viabiliza o emprego de técnicas que possibilitam a identificação de genótipos superiores capazes de gerar progênies com as características desejadas.
A identificação da divergência genética para a seleção de genótipos superiores pode ser realizada de várias maneiras, dentre elas destaca-se as técnicas biométricas, podendo ser utilizada de forma quantitativa ou preditiva. Na primeira temos como exemplo a análise dialélica, na qual é necessário realizar o cruzamento entre os genitores e posteriormente avaliar todas as combinações híbridas. No entanto, a necessidade de avaliações de p genitores e de todas suas combinações híbridas p(p-1)/2, pode se tornar onerosa e de difícil execução, principalmente quando o valor de p é elevado (Cruz et al., 2014).
Já a segunda temos os métodos preditivos da divergência entre genitores, que dispensam a obtenção prévia das combinações hibridas devido considerar as diferenças agronômicas, morfológicas, fisiológicas ou moleculares, entre outras, exibidas pelos genitores na determinação da divergência genética (Cruz et al., 2014).
A escolha de genitores de feijão comum por meio de modelos preditivos com base na distância genética tem sido relatada por diversos autores (Barelli, et al., 2009; Stähelin et al., 2011; Leite et al., 2015; Santos et al., 2015; Sebim et al., 2016). Segundo Barili et al. (2011) a seleção de genitores contrastantes aumenta a possibilidade de gerar combinações híbridas superiores.
Neste contexto, este trabalho tem por objetivo avaliar a divergência genética entre genótipos de feijão comum e sugerir combinações híbridas contrastantes para produzir recombinações superiores.
A presente pesquisa foi realizada na área experimental do Centro de Pesquisa e Extensão da Empresa Mato-grossense de Pesquisa, Assistência e Extensão Rural (EMPAER), localizada no município de Cáceres-MT (16°13'42”S e 57°40'05”W), com altitude de 118 m. O solo é classificado como Argissolo Vermelho Amarelo Eutrófico Chernossólico (Arantes et al., 2012). Segundo a classificação de Köpeen, a região apresenta clima tropical quente e úmido (Awa), com duas estações bem distintas, período de chuva de outubro a março e regime de seca de abril a novembro (Neves et al., 2011).
Foram avaliados 21 genótipos de feijoeiro de diferentes grupos comerciais, sendo nove do grupo Carioca (BRS Estilo, BRS Horizonte, BRS Requinte, BRS 9435 Cometa, BRS Pontal, BRS Pérola, VC6, BRS MG Majestoso e IPR Tangará) cinco do grupo Preto (BRS Campeiro, BRS Valente, BRS Esplendor, BRS Grafite e BRS Supremo) dois do grupo Roxo (CNFRX 10531 e BRS Timbó) dois Manteigão tipo rajado (Iraí e BRS Radiante) e um do grupo dos seguinte grupos: Manteigão tipo jalo (Jalo Precoce), Rosinha (BRS Vereda) e Roxinho (BRS Pitanga). Excetuando o genótipo IPR Tangará que foi cedido pelo Instituto Agronômico do Paraná (IAPAR), todos os demais procedem do banco de Germoplasma do Centro Nacional de Pesquisa Arroz e Feijão (CNPAF/EMBRAPA).
O delineamento experimental adotado foi o de blocos casualizados, com três repetições. As parcelas foram compostas por quatro fileiras de plantas, com 4 m de comprimento, e espaçamento de 0,50 m entre si. A área útil foi formada pelas duas fileiras centrais. A semeadura foi realizada de forma convencional no dia 09 de maio de 2008, adotando-se a densidade de 12 plantas por metro linear. Previamente a semeadura, foi realizada adubação baseada na análise química do solo, com aplicação de 20, 90 e 48 Kg/ha-1 de N, P2O5 e K2O, respectivamente. Devido a época de plantio caracterizar-se como irrigado no Estado de Mato Grosso, foi utilizado o sistema de irrigação para evitar o déficit hídrico. Os tratos culturais foram realizados de maneira que a cultura não sofresse competição de plantas daninhas, pragas e doenças.
Foram avaliados os seguintes caracteres agronômicos: altura média da planta (ALTP) expressa em cm, obtida pela medição, da base do solo até o ápice da planta, em uma amostra aleatória de cinco plantas por parcela; diâmetro médio de caule (DMC) expresso em cm, obtido pela medição da base do caule, em uma amostra aleatória de cinco plantas por parcela; altura média da inserção da primeira vagem (ALTINSV) expressa em cm, obtido pela medição da base do solo até a inserção da primeira vagem, de uma amostra aleatória de cinco plantas por parcela; comprimento longitudinal médio das vagens (CLMV) expresso em cm, obtido pela medição, de uma extremidade longitudinal a outra da vagem, em uma amostra aleatória de cinco vagens, (imaturas, mas completamente desenvolvidas) por parcela; e peso médio de sementes (PMS), expresso em gramas, obtido pela pesagem da amostra de 100 grãos por parcela.
Os dados obtidos para cada característica foram submetidos à análise de variância com a comparação das médias realizada pelo teste de Scott e Knott a 5 % de probabilidade, considerando o efeito do genótipo como fixo. Para a avaliação da variabilidade genética total existente entre os genótipos, foi utilizada análise multivariada, com base na distância generalizadas de Mahalanobis. No estudo de divergência genética, foram utilizados os métodos aglomerativos de otimização de Tocher e Hierárquico “UPGMA” e calculado o coeficiente de correlação cofenética (r) e análises de variáveis canônicas, fundamentados na distância generalizada de Mahalanobis como medida de dissimilaridade. Todas as análises foram realizadas utilizando os recursos computacionais do software Genes (Cruz, 2013).
A tabela 1 apresenta o resumo das análises da variância dos caracteres agronômicos avaliados nos 21 genótipos de feijão comum, onde todos os caracteres avaliados apresentaram diferença significativa entre os genótipos ao nível de 1 % de probabilidade pelo teste F, excetuando a características ALTINSV que apresentou significância ao nível de 5 % de probabilidade. Estes resultados, demostram a existência da variabilidade entre os genótipos, sugerindo que tais materiais podem ser explorados quanto a seleção de genitores para hibridação objetivando ganhos genéticos em programas de melhoramento. Como apontando por Cruz et al. (2014), a existência de variabilidade em uma população é fator determinante para o sucesso em qualquer programa de melhoramento.
Tabela 1. Resumo da análise de variância, com base na média das cinco características
agronômicas avaliadas, nos 21 genótipos de feijoeiro, no município de Cáceres-Mato Grosso, 2008.

Os valores do coeficiente de variação experimental da análise de variância conjunta oscilaram de 7,51% (Característica ALTP) a 17,14 % (característica ALTINSV) (Tabela 1), o que de acordo com Pimentel Gomes (1985), estes coeficientes de variação são considerados baixos (inferiores a 10 %) e médios (10 a 20 %). Desta maneira, esses resultados indicam boa precisão experimental em relação a todos os caracteres avaliados na presente pesquisa. Resultados semelhantes para os coeficientes de variação aos encontrados nesta pesquisa foram obtidos por Coelho et al. (2010), para as características DMC, ALTINSV e CLMV e por Leite et al. (2015), para as características ALTP e PMS.
As estimativas de alguns parâmetros genéticos, são importantes para a definição das estratégias de melhoramento e para a identificação de genótipos superiores em relação às características que se pretende melhorar. Percebe-se que para todos os caracteres avaliados os valores estimados para a variância fenotípica (σ²F) foram próximos ao obtidos para a variação genética (σ²G) e ambos superiores aos valores observados para a variação ambiental (σ²E) (Tabela 1). Estes resultados indicam maior influência dos componentes genéticos quando comparados com o ambiental na expressão dos caracteres. Tal fato é confirmado pelas estimativas da herdabilidade com valores variando de 51.80% para ALTINSV a 91.29 % para PMS. Isso sugere a possibilidade de progresso genético com a seleção desses caracteres, uma vez que segundo Bueno (2006), a partir de 50 % se é possível realizar a manipulação da herdabilidade.
Fato relevante observado nesta presente pesquisa foi à magnitude do índice de variação (IV) superiores a 1,0 para os caracteres ALTP, CLMV e PMS. Este índice auxilia na detecção de variabilidade genética em uma população e representa a relação entre o coeficiente de variação genética e o coeficiente de variação experimental (Foltran et al., 1993). Segundo Vencovsky (1987) quando essa relação é igual ou maior que 1,0, a condição é altamente favorável para a seleção.
Os resultados para as médias dos 21 genótipos demostraram diferenças significativa ao nível de 5% para as características ALTP, DMC, CLMV E PMS, apenas a característica ALTINSV não apresentou diferença estatística (Tabela 2). Resultados semelhantes foram observados por Bonett et al. (2006) na avaliação de 58 acessos crioulas de feijão e cinco cultivares, por 11 caracteres morfoagronômicos, em que apenas a característica ALTINSV não apresentou diferença estatística.
Em relação a característica ALTP, os melhores valores ficaram entre 58,07 a 67,00 cm, sendo obtidos pelos seguintes genótipos (BRS Estilo, BRS Horizonte, BRS Vereda, CNFRX 10531, BRS Valente, BRS Timbó, BRS Requinte, BRS 9435 Cometa, BRS Esplendor, Pérola, BRS Grafite, VC 6, BRS Supremo, BRS Majestoso, IPR Tangará), em que os mesmos não apresentaram diferenças estatísticas entre si. Em pesquisa realizada por Salgado et al. (2011) avaliando o comportamento de cultivares de feijão comum no período da entressafra na região sul do Estado do Tocantins as cultivares BRS Esplendor, BRS Majestoso e a linhagem VC 6 apresentaram altura superiores com 71,55, 78,66 e 85,55 cm, respectivamente. De acordo com Simone et al. (1992) valores ideais para colheita mecanizada é acima de 50 cm, desta maneira todos os genótipos avaliados neste estudo apresentam valores satisfatórios para esta característica.
Para a característica DMC os maiores valores médios foram obtidos pelos genótipos (BRS Estilo, BRS Horizonte, BRS Vereda, BRS Valente, Iraí, BRS Requinte, BRS 9435 Cometa, BRS Esplendor, BRS Pitanga, Pérola, BRS Grafite, VC 6, BRS Supremo, BRS Majestoso, IPR Tangará) com variação entre 0,77 a 0,93 cm (Tabela 2). Esses resultados são superiores ao obtidos por Rocha et al. (2009), com a cultivar Pérola (0,68 cm) nos anos agrícolas de 2006/07 e 2007/08 em pesquisa realizadas em Lages, SC e por Oliveira et al. (2012) avaliando o potencial produtivo de genótipos de feijão comum em função do estresse de fósforo no Estado do Tocantins para a cultivar BRS-Requinte (0,46 cm).
Quanto à altura de inserção da primeira vagem os genótipos apresentaram valores variando entre 15,23 a 29,43 cm e não apresentaram diferenças significativas entre eles (Tabela 2). Oliveira et al. (2014) destaca que os valores para esta característica são considerados satisfatórios quando estão acima de 12 cm, pois facilita a colheita mecanizada, neste sentido, todos os genótipos avaliados nesta pesquisa apresentam altura de inserção da primeira vagem aceitáveis.
Para a característica comprimento longitudinal médio das vagens, os genótipos Jalo Precoce, Iraí e BRS Radiante, foram os superiores com valores de 13,13, 12,33 e 12,29 cm, respectivamente (Tabela 2). Os mesmos genótipos supracitados apresentaram também maior peso médio de sementes com 32,95, 33,87 e 32,80 g, respectivamente. Resultados semelhantes foram obtidos por Leite et al. (2015), em que estes genótipos estão entre os que se sobressaíram para esta característica e por Santos et al. (2011) com os genótipos Iraí e BRS Radiante.
Tabela 2. Análise comparativa de médias dos 21 genótipos de feijoeiro em relação
às cinco características avaliadas, Cáceres-Mato Grosso, 2008.
Média1/ das características avaliadas2/ |
|||||
Genótipos3/ |
ALTP |
DMC |
ALTINSV |
CLMV |
PMS |
1 |
62,27 a |
0,91 a |
19,07 a |
9,62 c |
19,32 c |
2 |
66,00 a |
0,81 a |
18,73 a |
11,02 b |
20,58 b |
3 |
62,67 a |
0,82 a |
29,43 a |
10,76 b |
19,17 c |
4 |
54,27 b |
0,70 b |
21,17 a |
10,07 c |
21,88 b |
5 |
59,07 a |
0,73 b |
21,18 a |
10,51 b |
21,30 b |
6 |
62,73 a |
0,93 a |
21,33 a |
9,44 c |
18,45 c |
7 |
58,07 a |
0,70 b |
20,57 a |
10,16 c |
17,67 c |
8 |
49,07 b |
0,77 a |
20,20 a |
12,29 a |
32,95 a |
9 |
48,13 b |
0,71 b |
15,90 a |
13,13 a |
33,87 a |
10 |
62,80 a |
0,80 a |
15,23 a |
10,51 b |
17,68 c |
11 |
66,80 a |
0,83 a |
17,00 a |
10,65 b |
19,33 c |
12 |
60,73 a |
0,79 a |
20,50 a |
9,79 c |
15,83 c |
13 |
54,60 b |
0,67 b |
18,67 a |
10,18 c |
19,20 c |
14 |
53,53 b |
0,65 b |
21,83 a |
12,33 a |
32,80 a |
15 |
49,67 b |
0,83 a |
18,87 a |
9,46 c |
17,32 c |
16 |
64,40 a |
0,82 a |
18,07 a |
11,11 b |
22,80 b |
17 |
62,80 a |
0,85 a |
20,83 a |
10,42 b |
22,37 b |
18 |
60,00 a |
0,83 a |
21,00 a |
10,53 b |
21,23 b |
19 |
67,00 a |
0,84 a |
21,27 a |
9,89 c |
18,60 c |
20 |
58,87 a |
0,78 a |
18,33 a |
10,59 b |
20,75 b |
21 |
66,07 a |
0,84 a |
19,57 a |
11,11 b |
24,42 b |
1/Médias seguidas de mesma letra não diferem significativamente a 5% de probabilidade pelo teste Scott Knott.
2/ALTP = altura média da planta; DMC = diâmetro médio de caule; ALTINSV = altura de inserção da primeira vagem; CLMV = comprimento longitudinal médio das vagens; PMS = peso médio de sementes.
3/(1) BRS Estilo, (2) BRS Horizonte, (3) BRS Vereda, (4) BRS Campeiro, (5) CNFRX 10531, (6) BRS Valente, (7) BRS Timbó, (8) Iraí, (9) Jalo Precoce, (10) BRS Requinte, (11) BRS 9435 Cometa, (12) BRS Esplendor, (13) BRS Pontal, (14) BRS Radiante, (15) BRS Pitanga, (16) Pérola, (17) BRS Grafite, (18)VC 6, (19) BRS Supremo, (20) BRS Majestoso, (21) IPR Tangará.
Na análise do agrupamento dos genótipos pelo método de otimização de Tocher, fundamentado na matriz de dissimilaridade utilizando a distância generalizada de Mahalanobis (D2ii’) obteve-se a formação de seis grupos (Tabela 3). O grupo I apresentou o maior número de genótipos contendo 52,38 %, dos genótipos avaliados. Todos os genótipos deste grupo apresentaram altura de inserção da primeira vagem variando de 15,23 cm (BRS Requinte) a 21,27 cm (BRS Supremo) e comprimento longitudinal médio das vagens entre 9,89 cm (BRS Majestoso) e 11,11 cm (BRS 9435 Cometa e IPR Tangará). O grupo II englobou os genótipos com baixo peso de sementes com valores de 19,32, 18,45 e 15,83 g, sendo composto pelos genótipos BRS Estilo, BRS Valente e BRS Esplendor, respectivamente.
Tabela 3. Grupo de genótipos de feijoeiro com padrões similares, estabelecido pelo método
de Tocher, utilizando a Distância Generalizada de Mahalanobis (D2ii’) como medida de dissimilaridade,
Cáceres-Mato Grosso.
Group |
Genotypes¹ |
I |
17, 18, 20, 5, 16, 2, 21, 11, 10, 19 e 7 |
II |
1, 6 e 12 |
III |
4 e 13 |
IV |
8, 9 e 14 |
V |
3 |
VI |
15 |
¹/(1) BRS Estilo, (2) BRS Horizonte, (3) BRS Vereda, (4) BRS Campeiro, (5) CNFRX 10531, (6) BRS Valente,
(7) BRS Timbó, (8) Iraí, (9) Jalo Precoce, (10) BRS Requinte, (11) BRS 9435 Cometa, (12) BRS Esplendor,
(13) BRS Pontal, (14) BRS Radiante, (15) BRS Pitanga, (16) Pérola, (17) BRS Grafite, (18)VC 6,
(19) BRS Supremo, (20) BRS Majestoso, (21) IPR Tangará.
Já o grupo III foi formado pelos genótipos BRS Campeiro e BRS Pontal, tais genótipos apresentaram valores muitos próximos para as características DMC e CLMV com 0,70 cm e 0,67 cm para DMC e 10,07 cm e 10,18 cm para CLMV, respectivamente. No Grupo IV concentrou os genótipos de origem Andina, destacam-se no quesito similaridade quanto as características comprimento longitudinal médio das vagens e peso médio de sementes em que obtiveram às maiores médias dentro todos os genótipos avaliados.
Os grupos V e VI foram os menores grupos, sendo composto por apenas um genótipo BRS Vereda e BRS Pitanga, respectivamente. A separação destes genótipos deve-se ao fato do BRS Vereda apresentar altura de inserção da primeira vagem superior aos demais genótipos avaliados (29,43 cm) e o BRS Pitanga apresentou baixo CLMV (9,46 cm) e ALTP (49,67 cm).
De acordo com o dendrograma obtido pelo método hierárquico UPGMA (Figura 1), os genótipos foram reunidos em três grupos com subdivisões para o grupo I e II, sendo que o grupo I foi composto por quatro subdivisões, em que os genótipos BRS Grafite, VC 6, CNFRX 10531 e BRS Majestoso constituíram o subgrupo I-A, os genótipos BRS Timbó, BRS Pontal e BRS Campeiro o subgrupo I-B, os genótipos Pérola, IPR Tangará, BRS Horizonte, BRS 9435 Cometa e BRS Requinte o subgrupo I-C e, o genótipo BRS Vereda isolado no subgrupo I-D; o Grupo II composto por duas subdivisões, os genótipos BRS Estilo, BRS Valente, BRS Esplendor e BRS Supremo no subgrupo II-A, e o genótipo BRS Pitanga isolado no subgrupo II-B; o Grupo III composto pelos genótipos Iraí, BRS Radiante e Jalo Precoce. A alta correlação cofenética (0,91) mostra a consistência da análise de agrupamento em relação à matriz de dissimilaridade

Figura 1. Dendrograma ilustrativo da divergência genética entre 21 genótipos de feijoeiro,
obtido pelo método de ligações médias (UPGMA), utilizando a Distância Generalizada
de Mahalanobis (D2ii’) como medida de dissimilaridade. *R: Correlação cofenética.
Os métodos aglomerativos de otimização Tocher e Hierárquico UPGMA, foram parcialmente similares, ficando evidente a divisão dos grupos em relação ao grupo gênico de origem, pois em ambos os agrupamentos os genótipos de feijão de origem Andina estão em um grupo isolado em relação aos genótipos de origem Mesoamericana. A separação de genótipos de acordo com o centro de origem também foi evidenciada em pesquisas realizadas por Barelli et al. (2009). Os genótipos BRS Vereda e BRS Pitanga, que formavam grupos isolados pelo método de Tocher, quando realocados para grupos maiores pelo método UPGMA, formaram subgrupos isoladamente em seus respectivos grupos.
Em relação a divergência entre os métodos aglomerativos ocorreu a realocação dos genótipos do Grupo III pelo método de Tocher para o subgrupo I-B; realocação do genótipo BRS Supremo do Grupo I (Tocher) para o subgrupo II-A (UPGMA), os genótipos BRS Vereda (Grupo V - Tocher) e BRS Pitanga (Grupo VI - Tocher) deixaram de constituir grupos isolados, sendo realocados para o subgrupo II-B e subgrupo I-D (UPGMA), respectivamente.
Analisando a divergência genética com base nas variáveis canônicas (Tabela 4), verificou-se que as duas primeiras variáveis foram capazes de explicar 89,76% da variação total. Segundo Cruz et al. (2014), para que haja uma boa interpretação da variabilidade entre os genótipos é necessária que as duas primeiras variáveis canônicas permitam estimativas mínimas de 80% da variação total contida no conjunto de caracteres. Assim, conforme apontando pelos autores citados acima é possível explicar de maneira satisfatória e confiável a variabilidade dos genótipos da presente pesquisa em um gráfico no plano bidimensional com grau desprezível de distorção provocada pelas distâncias entre os genótipos. Resultados semelhantes foram observados por Barelli et al. (2009) em que as duas primeiras variáveis canônicas explicaram 81,90 % da variação total. Resultados diferentes foram obtidos por Cabral et al. (2011), em que se precisou de três variáveis canônicas para explicar 82,16 % da variação total.
Tabela 4. Autovalores (λi) correspondentes às porcentagens de variação, explicadas pelas Variáveis
Canônicas (VCi), e coeficientes de ponderação (autovetores) de cinco características, avaliadas em
21 genótipos de feijoeiro, Cáceres-Mato Grosso, 2008.
|
Autovalores |
|
Coeficientes de Ponderação Associados1 |
|||||
VCi |
λi |
λi % |
% |
ALTP |
DMC |
ALTINSV |
CLMV |
PMS |
VC1 |
11,7262 |
78,16 |
78,16 |
-0,7736 |
-1,2726 |
0,00761 |
1,40725 |
0,35662 |
VC2 |
1,7405 |
11,60 |
89,76 |
0,2222 |
-0,6153 |
-0,11827 |
1,0612 |
-0,04983 |
VC3 |
0,7674 |
5,11 |
94,87 |
-0,2108 |
11,6534 |
0,13391 |
-0,17747 |
0,11622 |
VC4 |
0,6546 |
4,36 |
99,23 |
0,06206 |
-6,3982 |
0,21251 |
0,1134 |
-0,1137 |
VC5 |
0,1149 |
0,77 |
100 |
-06103 |
3,41423 |
0,03238 |
0,97094 |
-0,18683 |
1/ALTP = altura média da planta; DMC = diâmetro médio de caule;
ALTINSV = altura de inserção da primeira vagem;
CLMV = comprimento longitudinal médio das vagens;
PMS = peso médio de sementes.
Os grupos formados por meio da dispersão gráfica no espaço bidimensional dos 21 genótipos, utilizando-se a primeira e a segunda variáveis canônicas (Figura 2), estão em acordo com os grupos obtidos pelos métodos de otimização de Tocher (Tabela 3) e método hierárquico UPGMA (Figura 1), em que é possível observar novamente a separação dos genótipos de acordo com grupo gênico Andino e Mesoamericano, e a similaridade entre alguns genótipos dentro destes dois grupos.
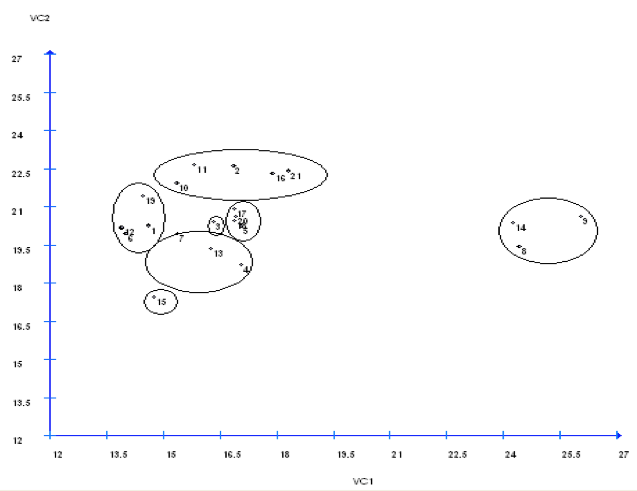
Figura 2. Dispersão gráfica dos escores, em relação aos dois eixos representativos das duas
primeiras variáveis canônicas (VC1 e VC2), obtidas de cinco características avaliadas
em 21 genótipos de feijoeiro, Cáceres-MT.
Baseando-se na análise de agrupamento das médias, juntamente com os agrupamentos estabelecidos pelo método de Tocher, UPGMA e variáveis canônicas, é possível inferir sobre quais serão os cruzamentos promissores e quais poderão resultar restrita variabilidade nas gerações segregantes. Segundo Barili et al. (2011), quanto maior o grau de divergência entre os genitores, maior será a variabilidade resultante na população segregante, e maior é a probabilidade de reagrupar os alelos em novas combinações favoráveis.
A obtenção de populações superiores é um dos principais objetivos dos programas de melhoramento, neste sentido, a escolha de genótipos com maior dissimilaridade e com médias elevadas para os caracteres avaliados, possibilita ter uma predição da obtenção de segregantes promissoras. As combinações que atendem a estes princípios dentro dos genótipos avaliados são: BRS Majestoso x Iraí, BRS Majestoso x Jalo Precoce, BRS Majestoso x BRS Radiante, BRS Grafite x Iraí, VC 6 x Jalo Precoce e BRS Horizonte x BRS Radiante.
Os genótipos avaliados apresentam variabilidade genética e potencial para o melhoramento genético.
populações segregantes promissoras podem ser obtidas com as hibridações entre os seguintes genótipos: “BRS Majestoso x Iraí ”, “BRS Majestoso x Jalo Precoce”, “BRS Majestoso x BRS Radiante”, “BRS Grafite x Iraí”, “VC 6 x Jalo Precoce” e “BRS Horizonte x BRS Radiante”.
Arantes, E. M.; Cremon, C.; y Luiz, M. A. C. (2012). Alterações dos atributos químicos do solo cultivado no sistema orgânico com plantio direto sob diferentes coberturas vegetais. Revista Agrarian, 5(15), p.47-54. Recuperado de http://ojs.ws.ufgd.edu.br/index.php?journal=agrarian&page=article&op=view&path%5B%5D=563&path%5B%5D=1012
Barelli, M. A. B.; Gonçalves-Vidigal, M. C.; Vidigal Filho, P. S.; Neves, L. G.; y Silva, H. T. da. (2009). Genetic divergence in common bean landrace cultivars from Mato Grosso do Sul State. Semina: Ciências Agrárias, 30 (supl.1), p.1061-1072. Recuperado de http://dx.doi.org/10.5433/1679-0359.2009v30n4Sup1p1061.
Barili, L. D.; Vale, N.M.; Arruda, B.; Toaldo, D.; Rocha, F.; Coimbra, J. L. M.; Bertoldo, J. G.; y Guidolin, A. F. (2011). Escolha de genitores contrastantes para compor o bloco de cruzamentos de genótipos de feijão. Revista Brasileira de Agrociência, 17(3-4), p.303-310. Recuperado de http://dx.doi.org/10.18539/CAST.V17I3.2063.
Bonett, L. P.; Gonçalves-Vidigal, M. C.; Schuelter, A. R.; Vidigal Filho, P. S.; Gonela, A. y Lacanallo, G. F. (2006). Divergência genética em germoplasma de feijoeiro comum coletado no estado do Paraná, Brasil. Semina: Ciências Agrárias, 27(4), p.547-560. Recuperado de http://dx.doi.org/10.5433/1679-0359.2006v27n4p547.
Bueno, L. C. S. (2006). Melhoramento genético de plantas. Princípios e procedimentos. Lavras: Universidade Federal de Lavras, 2006, 319 p.
Cabral, P. D. S.; Soares, T. C. B.; Lima, A. B. P.; Alves, D. S.; y Nunes, J. A. (2011). Diversidade genética de acessos de feijão comum por caracteres Agronômicos. Revista Ciência Agronômica, 42(4), p. 898-905. Recuperado de http://www.ccarevista.ufc.br/seer/index.php/ccarevista/article/view/1277/624.
Coelho, C.M.M.; Zilio, M.; Souza, C.A.; Guidolin, A.F.; y Miquelluti, D. J. (2010). Características morfo-agronômicas de cultivares crioulas de feijão comum em dois anos de cultivo. Semina: Ciências Agrárias, 31(4) supl.1, p. 1177-1186. Recuperado de http://dx.doi.org/10.5433/1679-0359.2010v31n4Sup1p1177.
Cruz, C. D. Carneiro, P. C. S.; Regazzi, A. J. (2014). Modelos Biométricos Aplicados ao Melhoramento Genético. v. 2, 3ª ed. Viçosa: Editora UFV, Universidade Federal de Viçosa. 668p.
Cruz, C.D. (2013). GENES - a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum, 35(3), p.271-276. Recuperado de http://dx.doi.org/10.4025/actasciagron.v35i3.21251.
Foltran, D. E.; Gonçalves, P. S.; Sabino, J. C.; Igue, T.; y Vilela, R. C. F. (1993). Estimativas de parâmetros genéticos e fenotípicos em mamão. Bragantia, 52 (1), p.7-15. Recuperado de http://dx.doi.org/10.1590/S0006-87051993000100002
Leite; P, H, M.P.; Barelli; M, A. A.; Neves, L. G.; Silva; C. R, da.; y Oliveira, T. C. (2015). Genetic divergence among common bean cultivars from precocious group. Científica, 43 (2), p.143-148. Recuperado de http://dx.doi.org/10.15361/1984-5529.2015v43n2p143-148.
Neves, S. M. A. da.; Nunes, M. C. M.; y Neves, R. J. (2011). Caracterização das condições climáticas de Cáceres/MT-Brasil, no período de 1971 a 2009: subsídio às atividades agropecuárias e turísticas municipais. Boletim Goiano de Geografia, 31 (2), p. 55-68, Recuperado de http://dx.doi.org/10.5216/bgg.v31i2.16845.
Oliveira, A. C.; Silva, J.; Santos, M. M.; Cancellier, E. L.; y Fidelies, R, R. (2014). Desempenho agronômico de cultivares de feijão em função da adubação fosfatada no sul do Estado do Tocantins. Revista Caatinga, 27(1), p. 50–59. Recuperado de http://periodicos.ufersa.edu.br/revistas/index.php/sistema/article/view/2657/pdf_85.
Oliveira, T. C. de.; Silva, J.; Sousa, S. de A.; Campestrini, R.; Fidelis, R. R. (2012). Potencial produtivo de genótipos de feijão comum em função do estresse de fósforo no Estado do Tocantins. Journal of Biotechnology and Biodiversity, 3(3), p.24-30. Recuperado de http://revista.uft.edu.br/index.php/JBB/article/view/285.
Pimentel GOMES, F. (1985). Curso de estatística experimental. São Paulo: Nobel, 467 p.
Rocha, F. da.; Barili, L. D.; Garcia, S. H.; Modena, R.; Coimbra, J. L. M.; Guidolin, A. F.; y Bertoldo, J. G. (2009). Seleção em populações mutantes de feijão (Phaseolus vulgaris L.) para caracteres adaptativos. Biotemas, 22 (2), p.19-27. Recuperado de http://dx.doi.org/10.5007/2175-7925.2009v22n2p19.
Rocha, F. da.; Stinghen, J. C.; Gemeli, M. S.; Coimbra, J. L. M.; y Guidolin, A. F. (2014). Análise dialélica como ferramenta na seleção de genitores em feijão. Revista Ciência Agronômica, 45(1), p.74-81. Recuperado de http://www.ccarevista.ufc.br/seer/index.php/ccarevista/article/view/2040/905.
Salgado, F. H. M.; Fidelis, R. R.; Carvalho, G. de L.; Santos, R. dos S.; Canceillier, E. L.; y Silva, G. F. (2011). Comportamento de genótipos de feijão, no período da entressafra, no sul do estado de Tocantins. Bioscience Journal. 27(1), p.52-58.
Santos, A. dos.; Braga, D. C.; Correa, A. M.; Melo, C. L. P. de.; Silva, J. A. dos.; Lima, A. R. S.; y Rodrigues, E. V. (2015). Escolha de genitores de feijão-comum baseado na divergência genética. Revista Agrarian, 8(29), p.235-245.
Santos, A.; Correa, A. M.; Melo, C. L. P.; Durante, L. G. Y.; Carneiro, T.; y Oliveira, R. (2011). Desempenho agronômico de genótipos de feijão comum cultivados no período “da seca” em Aquidauana-MS. Revista Agrarian, 4(11) p.33-42. Recuperado de http://ojs.ws.ufgd.edu.br/index.php?journal=agrarian&page=article&op=view&path%5B%5D=839&path%5B%5D=674.
Sebim, D.E.; Oliveira, P. H.; Brusamarello, A. P.; Baretta, D. R.; Brum, B. (2016). Diversidade genética entre populações de feijão crioulo através da análise multivariada de caracteres morfoagronômicos. Espacios, 37(16) p.19. Recuperado de http://www.revistaespacios.com/a16v37n16/16371619.html.
Simone, M. de; Failde, V.; Garcia, S.; y Panadero, P.C. (1992). Adaptación de variedades y líneas de judías secas (Phaseolus vulgaris L.) a la recolección mecanica directa. Salta : INTA, 5p.
Singh, S. P.; Gepts, P.; y Debouck, D. B. Races of common bean (Phaseolus vulgaris, Fabaceae). Economic Botany, 45(3) p.379-396, 1991. Recuperado de http://dx.doi.org/10.1007/BF02887079.
Stähelin, D.; Valentini, G.; Andrade, L. R. B.; Verissimo, M. A. A.; Bertoldo, J. G.; Guidolin, A. F.; y Coimbra, J. L. M. (2011). Screening multivariado entre acessos e cultivares de feijão do grupo preto para utilização em blocos de cruzamento. Biotemas, 24(1), p.95-103.
Tsutsumi, C. Y.; Bulegon, L. G.; y Piano, J.T. (2015). Melhoramento genético do feijoeiro: avanços, perspectivas e novos estudos, no âmbito nacional. Nativa, 3(3), p.217-223.
Vencovsky, R. (1987). Herança quantitativa. In: PATERNIANI, E. & VIÉGAS, G.P., coords. Melhoramento e produção do milho no Brasil. 2.ed. Campinas, Fundação Cargill, cap. 5, p.137-214.
1. Engenheiro Agrônomo, Universidade do Estado de Mato Grosso – UNEMAT, Cáceres-MT; e-mail: leosulzbacher@gmail.com
2. Doutorando do Programa de Pós-Graduação em Biodiversidade e Biotecnologia da Rede BIONORTE; e-mail: silvabiologo@hotmail.com
3. Doutorando do Programa de Pós-Graduação em Biodiversidade e Biotecnologia da Rede BIONORTE; e-mail: brunowzago@hotmail.com
4. Pós Doutoranda pelo Programa de Programa de Pós-Graduação em Genética e Melhoramento de Plantas, UNEMAT - Universidade do Estado de Mato Grosso, Cáceres, MT; e-mail: correa.carla@yahoo.com.br
5. Doutoranda do Programa de Pós-Graduação em Biodiversidade e Biotecnologia da Rede BIONORTE; E-mail: melao.a.v.bio@gmail.com
6. Professor Dr. Adjunto, Departamento de Agronomia, Universidade do Estado de Mato Grosso - UNEMAT, Cáceres-MT; e-mail: mbarelli@unemat.br